








































モニタリング活動
植生調査 2013年度報告書
1.はじめに
富士山南麓では1996年(平成8年)9月の台風17号により大規模な風倒被害が発生し,壮齢ヒノキ林を中心に約90,000m3が被害を受けた.富士山「まなびの森」事業計画では,この風倒被害跡地に広葉樹を中心とした在来樹種を植栽することによって,早期に自然林を復元することを目指している.
本調査は,「まなびの森」区域内での台風による風倒跡地の森林の回復および変化について,次の3つの視点からモニタリングをおこない,自然林の早期再生のための管理・施業方法に関する情報を得ようとするものである.
1)風倒被害跡地の群状植栽地の森林回復過程について,植栽木の生育状況,群落構造の発達,林床を含めた種組成の変化を中心にモニタリングをおこなう.同時に区域内のブナ自然林,ヒノキおよびウラジロモミ人工林についても同じ項目をモニタリングし,これらとの比較から群状植栽地の回復状態を評価する.
2)風倒跡地に樹木を植栽しない場合の植生変化のモニタリングをおこない,天然更新の可能性を探索する.また調査地周辺にはニホンジカが生息するため,シカの採食圧が森林の回復にどの程度影響するかを明らかにする.
3)隣接ヒノキ林の風倒にともなうブナ自然林の質的変化をモニタリングし,風倒が周辺域に及ぼす二次的影響について明らかにする.
今年度の調査では上記の目的1)に関して,2000年から2001年にかけて群状植栽地に設置した固定調査区2ヶ所の追跡調査をおこなった.この結果を前回までの調査結果と比較することによって,これまでの植生変化および森林の再生状況を明らかにした.また,「まなびの森」区域内では,シカの採食圧の増加にともなって,バイケイソウなどの不嗜好植物の繁茂をはじめとした植生の変化や,土壌の流出が顕著になってきた.そこで,目的2)3)に関して,ブナ自然林の一部に防鹿柵を設置し,柵の内外に新たな固定調査区を設けて,柵の設置による林床植生保護の効果を検証するための調査を開始した.
2.調査区の概要
「まなびの森」区域内の風倒被害跡地では,1998年から2002年にかけて,周辺の自然林に生育する樹種の植栽がおこなわれている.ここでは,風倒木の伐採・搬出後,一般的な地拵えはおこなわず,伐根などの残材を集積・放置してある.この放置枝条の間に,ひとつの樹種を約20m四方の群状に植栽した(群状植栽).また,基線上500-550m付近の約1haの範囲では,風倒木の搬出後,植栽をおこなわずに自然状態での森林再生を試みている.
本区域内に,森林の再生状況をモニタリングするための固定調査区を9ヶ所設置し(図1),2000年以降モニタリング調査をおこなってきた.今回,調査対象としたのはケヤキ植栽地とブナ植栽地の固定調査区である.各調査区におけるこれまでの調査実施年を表1に示す.
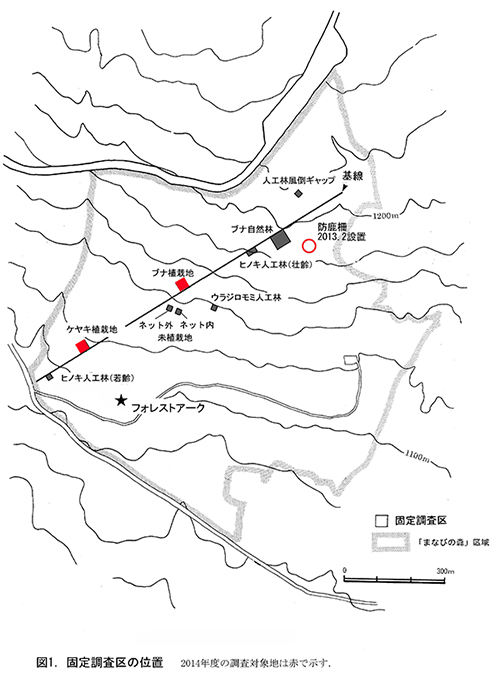
①ケヤキ植栽地
1999年に植栽がおこなわれた場所に,翌2000年に設置した調査区であり,植栽後14年が経過している.面積は20m×20mの400㎡で,10m×10mの区画に4区分されている.調査区内には主としてケヤキが植栽されているが,コナラやイロハモミジも少数含まれている.ここでは,2000年に毎木調査,2001年に種組成の調査がおこなわれ,2003年,2007年,2010年に同じ項目が再度測定された.今回の調査は4度目の追跡調査となる.
②ブナ植栽地
2001年に植栽がおこなわれた場所に,植栽直後の2001年に設置された調査区であり,植栽後12年が経過している.ケヤキ植栽地と同様,面積は20m×20mの400㎡で,10m×10mの区画に4区分されている.調査区内には主にブナが植栽されている.ここでは,2001年に毎木調査,2002年に種組成の調査がおこなわれ,2004年,2007年,2010年に同じ項目が再度測定された.今回の調査は4度目の追跡調査となる.
表1.各固定調査区の調査実施年
*種組成の調査のみ実施.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
③防鹿柵内外の調査区
また, 2013年3月に,ブナ自然林の固定調査区から約50m東側の緩斜面に,およそ13m×13m程度の防鹿柵を設置した(図1).ネットはステン線入りナイロン製,目開き径5cmで,高さ2mで支柱間隔は約3mである.この内部に10m×10mの固定調査区を設置した.また,防鹿柵の効果を検証するための比較対象として,柵に近接する場所に同じサイズの対照区を設置した.
3.調査方法
(1)植栽地の回復状況に関する調査
①毎木調査
ブナ植栽地,ケヤキ植栽地のそれぞれで,調査区内に生育している樹木について,植栽・非植栽にかかわらず,各個体の樹高および根元直径を測定した.高木性樹種については樹高30cm以上,低木性樹種については樹高130cmの個体を調査対象とした.樹高は検測棹をもちいてcm単位で,根元直径はノギスを用いて㎜単位で計測した.また,個体識別のためにナンバーテープをつけ,位置図を作成した.
②植生調査
10m×10mの区画を単位として,植物社会学的な方法による植生調査をおこなった.これは,出現するすべての植物をリストアップして,それぞれの種の優占度および群度を判定するものである.優占度・群度はBraun-Blanquet(1964)の判定基準にもとづき,以下のように判定した.
優占度(種の個体数と被度を表す)
| r | : | 単独で生育 |
| + | : | まばらに生育し被度はごく小さい |
| 1 | : | 個体数は多いが被度は小さい(1/10以下) |
| 2 | : | 非常に個体数が多い,または被度が1/10~1/4 |
| 3 | : | 被度が1/4~1/2 |
| 4 | : | 被度が1/2~3/4 |
| 5 | : | 被度が3/4以上 |
群度(種の群がりぐあいを表す)
| 1 | : | 単独で生育 |
| 2 | : | 小群状または束状に生育 |
| 3 | : | 斑状またはクッション状に生育 |
| 4 | : | 大きな斑状,または穴のあいたカーペット状に生育 |
| 5 | : | 一面に生育 |
(2)防鹿柵の効果に関する調査
防鹿柵内外に設置した固定調査区において,今後のモニタリングの初期データとするために,上記と同じ方法で植物社会学的な植生調査を行った.林床植生は季節によっても変化するため,春季(6月1日)と秋季(10月1日)に2回の調査を行った.また,シカによる採食は,林床草本の開花率の低下をもたらすことが知られているため,草本種については開花の有無を記録した.
4.調査結果
(1) 植栽地の回復状況
(1)-1.ケヤキ植栽地
①樹木個体数と空間分布
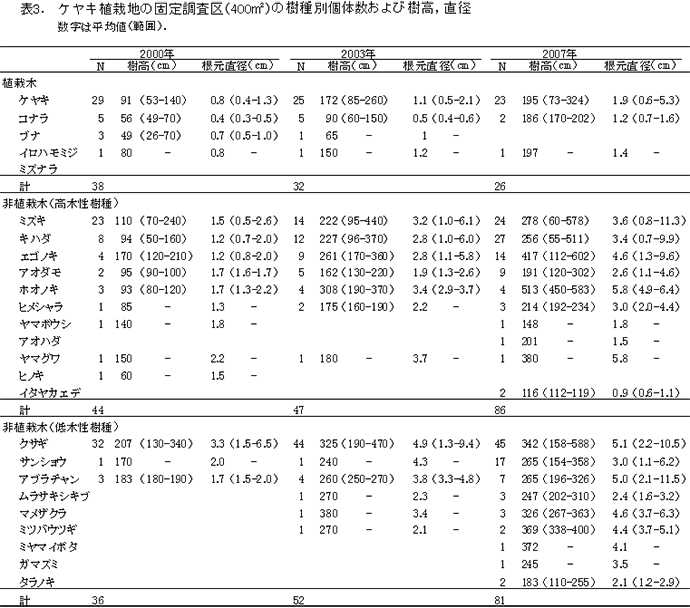
ケヤキ植栽地の固定調査区(400㎡)内に生育する樹木の樹種別の個体数と樹高・根元直径を表3に示した.また,図2にはそれらの分布を示した.
今回,測定対象となった樹木は135個体であった.新出木は1個体のみで,前回記録された個体のうち23個体が枯死していた.
生存していた植栽木は,ケヤキ19本,コナラ2本,イロハモミジ1本の計23本で,前回2010年の調査以降に枯死したのはケヤキ1本のみだった(表3).
非植栽木についてみると,高木性樹種では8種51本が生育していた(表3). 非植栽の高木性樹種の本数は,2007年の調査から減少に転じていたが,今回さらに本数を減らした.樹種別にみると,ミズキが2010年の15本から12本,キハダが16本から11本と減少し,13本で変わらなかったエゴノキが,初めて非植栽木で最も多い樹種となった.アオダモ(9本→7本)は微減,ホオノキ(3本)とヒメシャラ(3本)は変化がなかった.非植栽の高木性樹種に占める樹種別の割合は,ミズキ23.5%,キハダ21.6%,エゴノキ25.5%,アオダモ13.7%と前回とあまり変わっていない.前回まで記録されていたイタヤカエデとヤマグワは消失した.
低木性樹種では,8種62本の生育が記録された(表3).前回2010年の調査時から10本減少したが,枯死した10本のうち9本はクサギであった.クサギ以外の低木は,2007年以降ほとんど本数が変化していなかった.
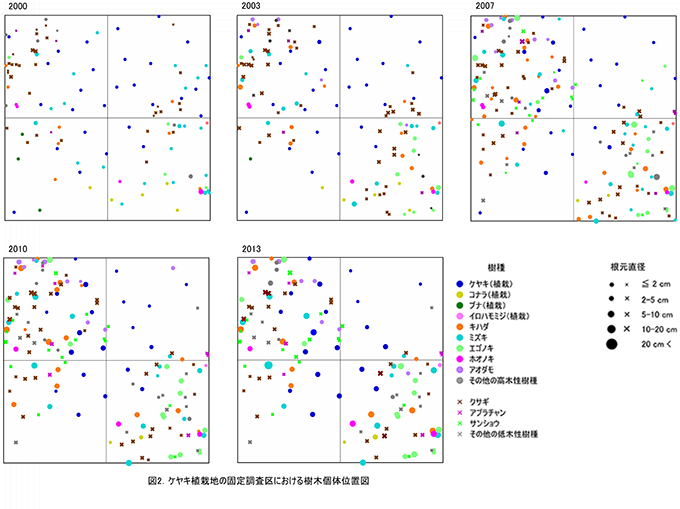
調査対象とした樹木の空間分布をみると,枝条が集積された区画ZP-2とZP-3への偏りが大きい(図2).これらの区画ではエゴノキ,ミズキ,キハダが根元直径10cm以上に成長している.一方で,それ以外の場所では植栽されたケヤキ以外の樹木はほとんど生育していなかった.過去3年間に枯死したクサギは,すべてこの区画に分布していたことから,放置枝条周辺で樹木が密生したことによる相互被陰が枯死の原因であると考えられる.
②樹木の成長と階層構造
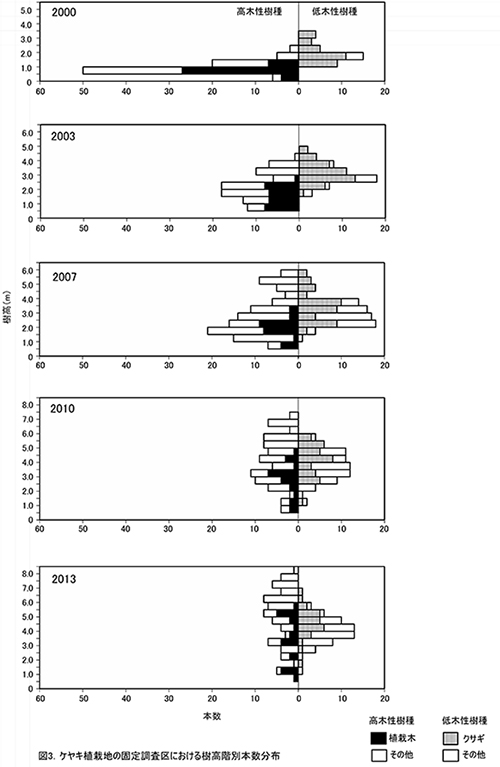
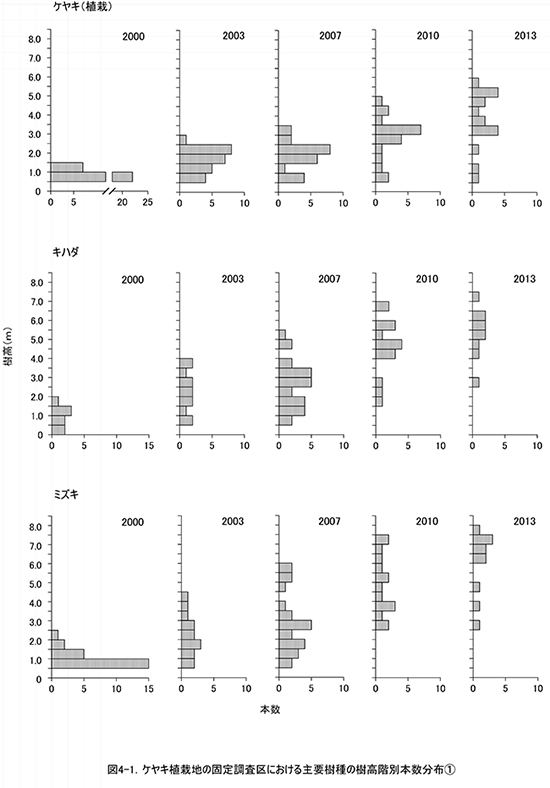
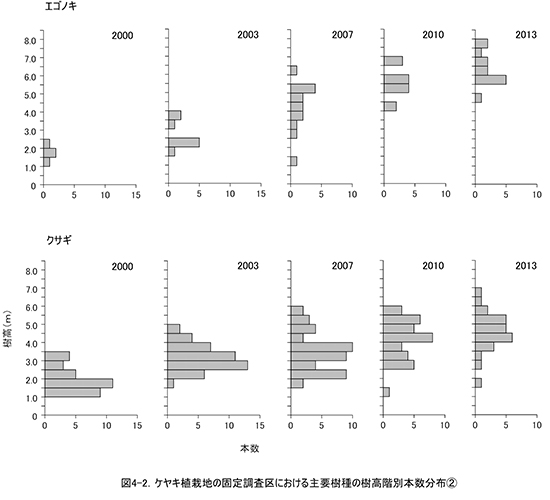
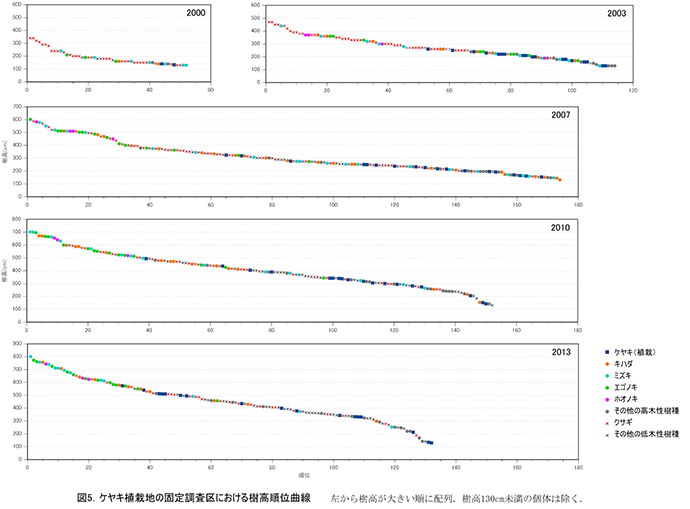
調査区内に生育する樹木の樹高階別本数分布の経年変化を図3に,そのうち個体数が多い樹種については図4に個別に示した.また,図5には1.3m以上の全個体の樹高を大きい順に並べた樹高順位曲線を示した.
全樹種の樹高階別本数分布の経年変化(図3)をみると,2000年には樹高1m未満のものが多かった高木性樹種の分布が,2003年から2007年にかけては二山形の分布になり,2010年以降はよりばらつきが大きくなっていることがわかる.二山形の分布が崩れたのは,高さ約3mから6m程度にクサギをはじめとする低木性樹種の階層が形成され,その下になった高木性樹種(特にミズキとキハダ)の多くが枯死したためである.2013年には,高さ6m以上の個体は,低木性樹種の階層を抜け出していることがわかる.植栽したケヤキの樹高は,まだ低木性樹種の高さを超えないものが多いが,位置的にほかの樹種と競合しない(図2)ため,枯死を免れている.
樹高順位曲線(図5)をみると,2010年から2013年にかけて,樹高5mを超える樹木の本数は40本程度で変わっていなかったが,樹高5mを超える部分の曲線の傾きが急になっていきている.このことは,低木性樹種が多い階層を抜け出した個体の成長速度が大きくなり,上位の階層を形成しつつあることを示している.樹高5m以上の上位40本のうち,クサギは4本にまで減少し,それ以外はほとんどがエゴノキ,ミズキ,キハダ,ホオノキであった.植栽したケヤキも5本が樹高5mを超え,クサギと同等以上に成長していた.なお,調査区内で最大の樹高をもつのは,ミズキの801cm,次いでエゴノキ773cm,762cm,ミズキとキハダ757cm,ホオノキ743cmと続いていた.
②樹木の成長と階層構造
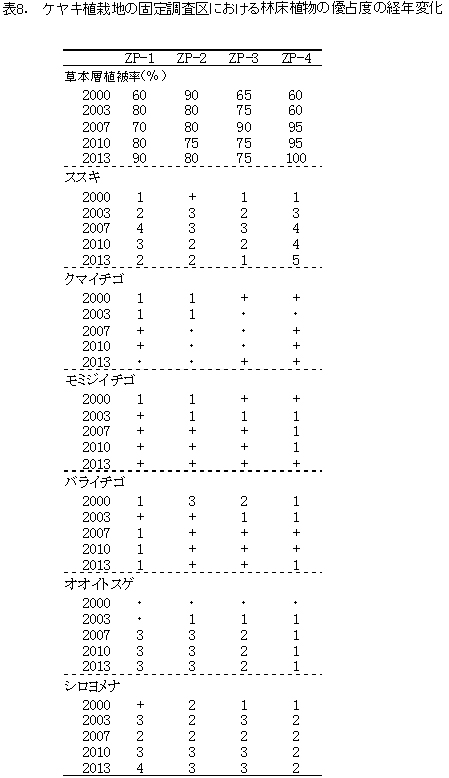
林床(草本層)に出現した植物のうち,林内の環境を反映する代表的なものについて,その優占度の変化を表8に示した.
現在の調査区の植生は,おおむね5mから8mの亜高木層,2mから5mの低木層,0.8m以下の草本層に区分できる.ただし,一部では低木層と草本層の分化は明瞭でない.区画ごとの出現種数は77種から99種であった.
亜高木層には,天然生の稚樹のうち生育のよいミズキ,キハダ,エゴノキ,ホオノキと,先駆性樹種のクサギが生育するほか,つる性木本のクマヤナギやツルウメモドキも出現した.低木層には,クサギ,ヤブウツギ,アブラチャン,サンショウ,コクサギ,ミツバウツギ,マメザクラ,ムラサキシキブなどが多く,亜高木層と同様の高木性樹種や植栽したケヤキも含まれていた.前回に比べてクサギが衰退したため,相対的にヤブウツギが多くなっていた.草本層では,ススキ,シロヨメナ,オオイトスゲが多く,それぞれ優占度1~5で出現していた.その他,組成的には前回と大きな変化はないが,本地域のブナ林に特徴的な草本種であるナベワリが新たに出現した.
次に草本層の主な種の優占度の変化(表8)をみると,ススキの優占度はZP1-3では減少してきているが,放置枝条がほとんどないZP-4では優占度5まで増加した.遷移初期に特徴的なキイチゴ属(クマイチゴ,モミジイチゴ,バライチゴ)は,遷移の進行にともなって消失すると予想されたが,まだ優占度は低いながら生育していた.林床生のスゲ属であるオオイトスゲは2007年以降ほとんど変化がない.また,シカの不嗜好植物であるシロヨメナは増加傾向にあり,ZP-1では優占度4でススキにかわる優占種となっていた.
(1)-2.ブナ植栽地
①樹木個体数と空間分布
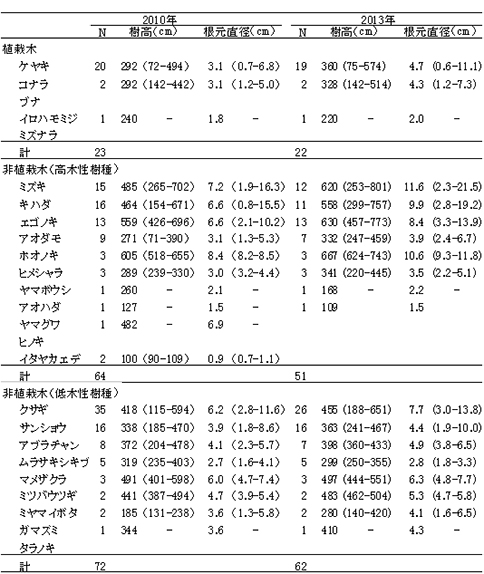
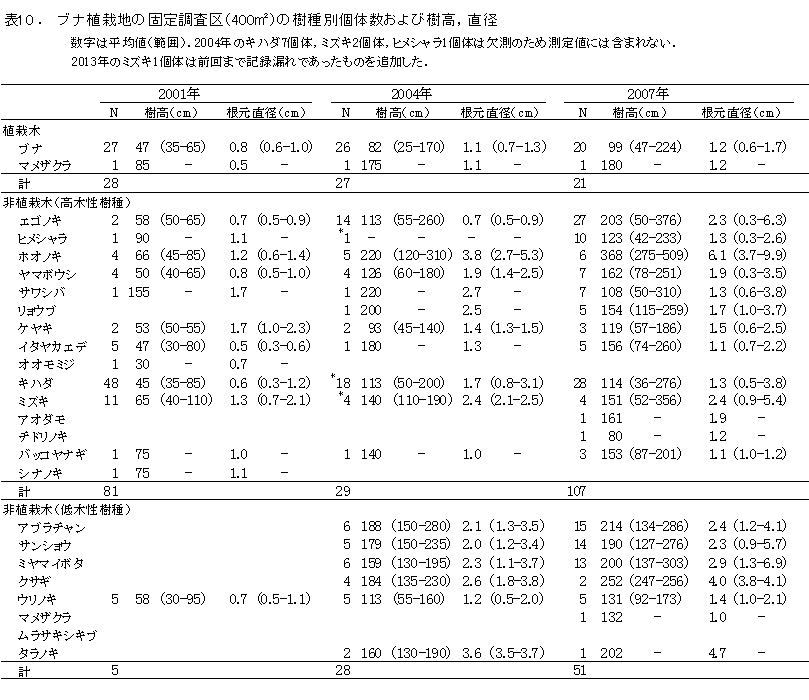
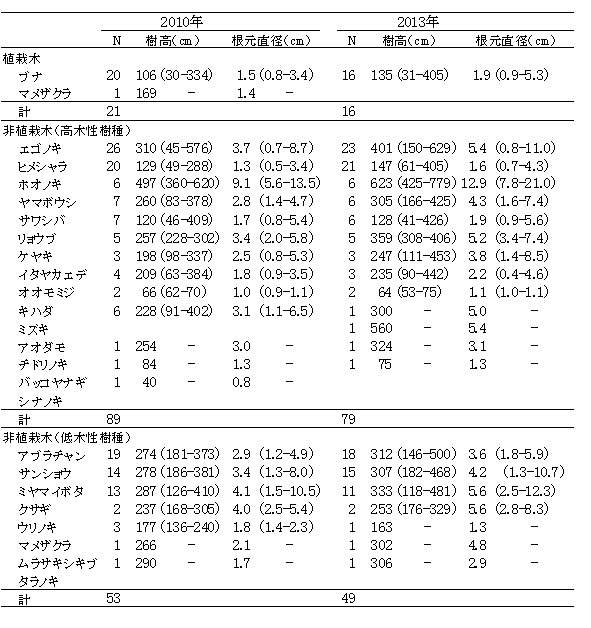
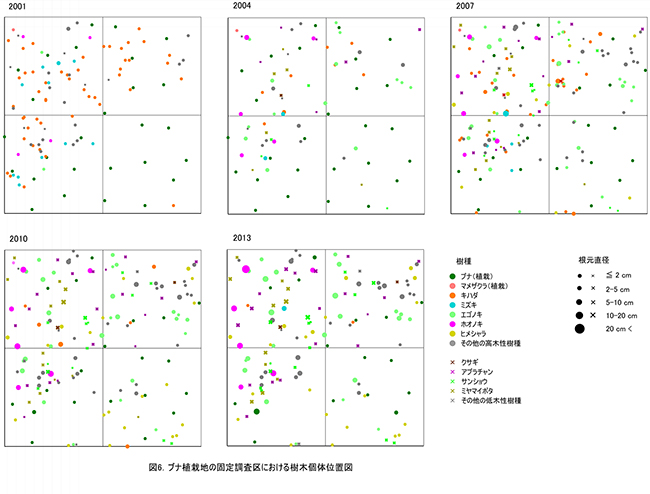
ブナ植栽地に設置した固定調査区(400㎡)内に生育する樹木の樹種別の個体数と樹高・根元直径を表10に示した.また,図6にはそれらの分布図を示した.
今回,測定対象となった樹木は144個体であった.そのうち新出個体は5個体,前回記録された個体のうち22個体が枯死していた.
植栽したブナは16本が生存しており,前回2010年の調査時から4本が枯死した.2007年から2010年には枯死個体はなかったが,この3年間で再び枯死が生じた.
非植栽木についてみると,高木性樹種は2010年の89本から10本減少して,13種79本が生育していた(表9).樹種別にみると,エゴノキが23本で,前回から3本減少したものの最も多く,ヒメシャラがそれに次ぐ21本であった.ホオノキ(6本→6本),ヤマボウシ(7本→6本),サワシバ(7本→6本),リョウブ(5本→5本)は前回と大きく変化はなかった.この調査区では,ケヤキ植栽地と異なりキハダ,ミズキの本数が少なく,キハダは前回より5本減少して1本,ミズキも1本しか生育していなかった.
低木性樹種は7種49本で,前回2010年の53本からわずかに減少した.個体数が多い樹種は,アブラチャン18本,サンショウ15本,ミヤマイボタが11本であった.ここではクサギの個体数は少なく,現在2本が生育するのみである.
調査対象とした樹木の空間分布をみると,樹木個体は放置枝条のある区画番号FP-1,FP-2,FP-4に多く,FP-3ではやや少ないことがわかる(図7). 放置枝条がある部分には,当初はキハダやミズキが多く生育していたが,現在ではエゴノキやホオノキが多くなっている.また,エゴノキやヒメシャラは,放置枝条がない区画にも生育しており,ススキが繁茂した中でも枯死せずに生育できることがわかる.
②樹木の成長と階層構造
調査区内に生育する樹木の樹高階別本数分布の経年変化を図7に,そのうち主要な樹種については図8に個別に示した.また,図9には1.3m以上の全個体の樹高を大きい順に並べた樹高順位曲線を示した.
全樹種の樹高階別本数分布の変化(図7)をみると,高木性樹種の個体数が最も多いのは0.5~1mの階級であり,ケヤキ植栽地と比べると樹高の小さい個体が多いことがわかる.これは,主にヒメシャラの実生が新たに加入しているためである.高木性樹種の成長のよい個体は5.0m以上に達し,2mから4m程度の低木性樹種の階層を超えていた.しかし,植栽木でこの階級に達しているものはなく,ブナの多くは依然として低木性樹種に被圧された状態にあるといえる.
樹高順位曲線の経年変化(図9)をみると,2013年にはホオノキ2個体が突出した樹高を示し,樹高5mを超える14本のうち12本までをホオノキとエゴノキが占めていた.その後には,サンショウ,ミヤマイボタ,アブラチャンなどの低木性樹種が続いており,植栽したブナは1本を除いてこれらの樹種よりも下位に位置していた.なお,上位の樹木の樹高は,ホオノキ779cm,690cm,635cm,635cm,エゴノキ629cm,606cmとなっていた.
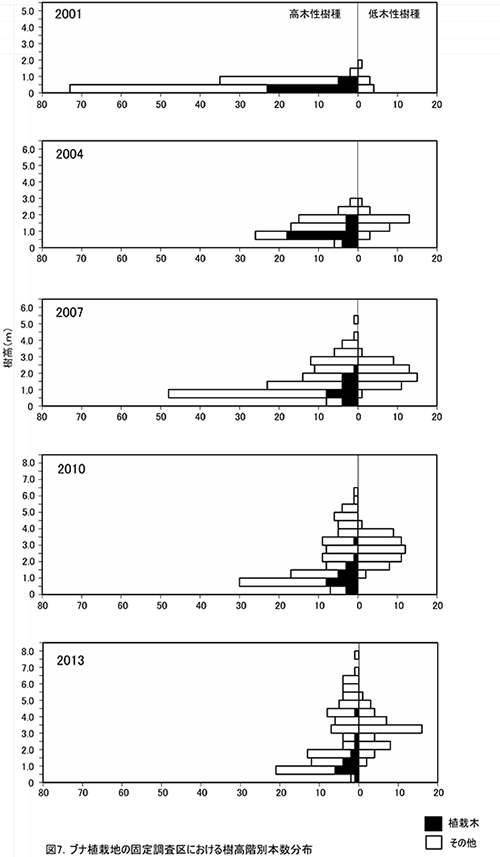
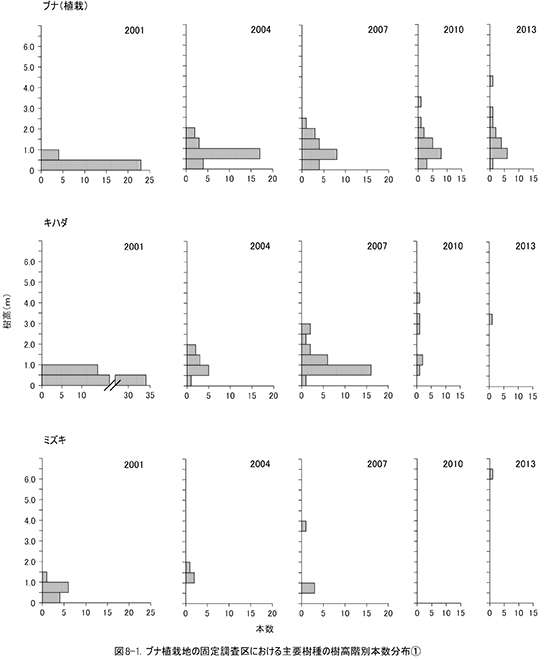
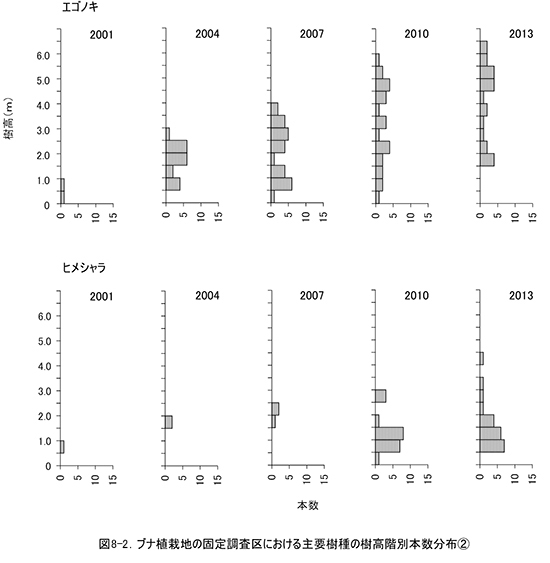
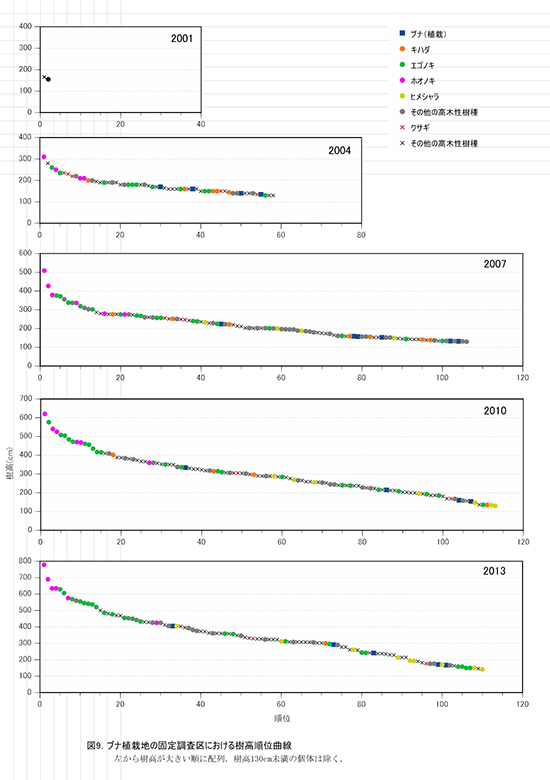
③林床植生の変化
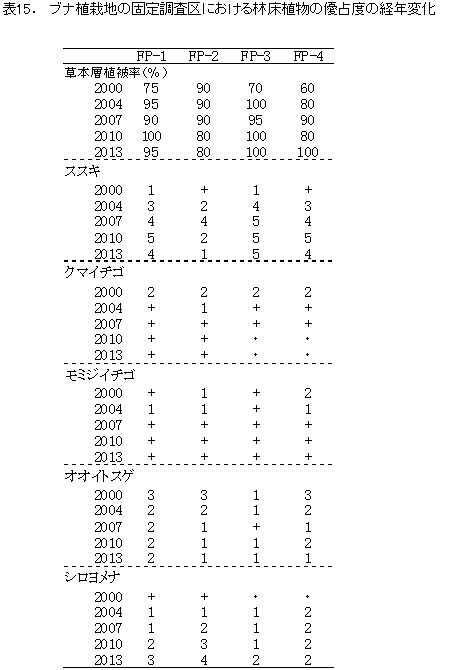
林床(草本層)に出現した植物のうち,林内の環境を反映する代表的なものについて,その優占度の変化を表15に示した.
現在の調査区内の植生は,一部で7m程度の亜高木層が形成されつつあるが,大部分は2mから5mの低木層と,2.0m以下の草本層に区分できる.草本層はススキが密生する階層に対応し,細かくみればその内部に1m以下の草本層第2層を認めることができるが,調査では区別しなかった.区画ごとの出現種数は75種から95種であった.
亜高木層にみられるのは,突出して成長しているホオノキとエゴノキのみである.低木層には,アブラチャン,サンショウ,ミヤマイボタ,ヤブウツギ,ウツギなどの低木性樹種や,エゴノキ,ヤマボウシなど高木性樹種の稚樹が生育している.ツルウメモドキ,サルナシ,サンカクヅルなどのつる性木本も出現する.草本層ではススキが優占しており,3区画で優占度4以上であった.ウツギ,ヤブウツギ,コアカソ,ヤマアジサイ,ミヤマイボタなどの低木も混生していた.シカが好まない不嗜好植物であるシロヨメナも多く生育していた.ケヤキ植栽地の調査区と同様,森林性のものから陽地性のものまで,さまざまな環境の種が混在していた.
草本層の主な種の優占度の変化(表15)をみると,2010年にはススキが3区画で優占度5となっていが,2013年にはそのうち2区画(FP-1,4)で優占度4に低下した.放置枝条の中心部で樹木の成長が旺盛な部分(FP-2)では2010年に既にススキの減少が観察されていたが,被圧により衰退がその周囲にも拡大してきたものと考えられる.地表撹乱後に生育しやすいクマイチゴやモミジイチゴは,2007年以降,優占度+に減少しているが,まだ消失してはいない.林床生のオオイトスゲも大きな変化はなかった.また,不嗜好植物であるシロヨメナは,4区画中3区画で優占度を増加させていた.特にススキの優占度が1まで低下した区画FP-2では,優占度4となっていた.
(1)-3. ケヤキ植栽地とブナ植栽地の回復状況の比較
2013年までの森林の回復状況は,ケヤキ植栽地とブナ植栽地の間で違いがみられる.ケヤキ植栽地では,樹高5m以上に成長した樹木が40本以上あり,その中心はミズキ,キハダ,エゴノキ,ホオノキであった.また,ススキは一部を除いて衰退しつつあり,樹林化が進んでいる.一方,ブナ植栽地では,樹高5m以上に成長した樹木は15本に満たず,そのほとんどはエゴノキとホオノキで,初期に多く発生していたミズキとキハダは著しく減少した.また,依然としてススキの優占度が高く,一部は草原状の相観を呈したままである.
このような違いは,植栽後の年数(ケヤキ植栽地のほうが2年早い)によるところも大きいが,放置枝条の量や斜面の傾斜といった,調査区間の立地の違いとも関係していると考えられる.
ケヤキ植栽地は,放置枝条の量が多く,置かれている面積も大きい.そのため,抜根した株の周囲に着いていた土壌中の埋土種子が早期から発芽し,多くの樹木が成長したと考えられる.また,ケヤキ植栽地は斜面傾斜が15度ほどの場所にあるため,放置枝条がある部分以外では,土壌の浸食が著しい.そのため,この部分ではそれほどススキが繁茂しなかったが,植栽したケヤキ以外の樹木も定着しにくかった.
一方,ブナ植栽地は,放置枝条の量が少なく,枝条が置かれた範囲でも地表面が見える程度であった.それに加えて,ケヤキ植栽地よりも緩傾斜であるため,土壌が流失しにくく,ススキが繁茂しやすい条件であった.そのため,当初は多く発生したキハダやミズキの実生は,ススキの優占度が最大になった2007年頃(表15)までに被陰されて枯死してしまった.それに変わって,初期成長は遅いが,ある程度被陰に耐性のあるエゴノキやヒメシャラの個体数が増加してきた.エゴノキやヒメシャラは,キハダやミズキが枝を横に広げて大型の葉を展開するのに対して,斜上する枝をもち葉も小さいので,ススキが密生した場所でも生育しやすいものと考えられる.
このように,ケヤキ植栽地とブナ植栽地では,その初期条件の違いによって,樹種構成がやや異なる樹林が形成される可能性が高い.ただし,どちらの場合も樹木の分布は放置枝条のある範囲に偏っており,パッチ状に樹林が形成されつつある.そのため,森林の再生は調査区内で均一に進むのではなく,まずパッチ状の樹林が形成され,それら樹冠が拡大することによって林冠が閉鎖されていくという過程をたどると予想される.
(2) 防鹿柵の効果
防鹿柵の内部に設置した調査区と,柵外に対照区として設置した調査区の林分は,いずれも高木層はブナの樹冠によって覆われ,シカの採食により高さ2m以下の低木層がほとんど消失していること,林床にはシカの不嗜性好植物を除くと小型の植物が広がっていることなど,構造的にも組成的もほぼ同質である.ただし,低木第1層の植被率は,柵内で50%,柵外で25%と差があった.柵外の調査区では,低木の枯死幹が多数みられ,低木層への採食の影響がより強く現れていた.以下に両調査区の概況を述べる.
(2)-1.防鹿柵内の固定調査区(調査区FN)
高木層はブナが優占し,やや樹高が低いオオモミジとミズキと合わせて植被率90%となる.亜高木層には,イトマキイタヤが生育し,調査区外に幹をもつヒメシャラ,ヤマボウシと合わせて植被率30%である.低木第1層は高さ2~6mと幅があり,イトマキイタヤ,サワシバの幼樹の他,アブラチャン,カマツカ,ミツバウツギなどの株立ちする低木が生育し,植被率50%となる.シカの採食の影響を受ける低木第2層はごくまばらで,高さ1.2mほど,植被率は3%である.わずかな萌芽枝が残るツクバネウツギやヤマウグイスカグラ,シカが食べないオニシバリなどが生育する.草本層には高さ0.1mほどの小型の草本が広がり,部分的に草丈の高い草本が0.3m程度の高さで生育している.部分的に植被がまばらな場所もあるため,植被率は65%である.小型の草本種としては,ツルシロカネソウ,セントウソウ,ヒメウワバミソウ,ヤマトグサ,イワボタン,コガネネコノメソウ,キクムグラなどが多い.高茎草本としては,アズマヤマアザミ,モミジガサ,カノツメソウ,ホウチャクソウなどが出現したが,モミジガサやホウチャクソウは小型化が著しく,開花個体はみられなかった.草丈が0.1mを超えているのは,主にシカ不嗜好性植物であるフタリシズカ,ヒロハノハネガヤ,バイケイソウ,ホソバテンナンショウなどであった.腐生植物のギンリョウソウや,寄生植物のヤマウツボの生育もみられた.また,人里の半陰地に多いアシボソ,ハナタデ,ミズヒキ,ムラサキケマンなども生育していた.
(2)-2.防鹿柵外の固定調査区(調査区FC,対照区)
高木層はブナが優占し,植被率は90%である.亜高木層には調査区内に生育する樹木はなく,調査区外に幹をもつサワシバ,ヤマボウシの樹冠がかかり,植被率20%である.低木第1層は高さ3~6mで,アズキナシ,サワシバの幼樹,アブラチャンが生育していた.植被率は防鹿柵内の調査区よりも低く25%であった.高さ1.0mの低木第2層はごくまばらで,コクサギ,ツクバネウツギの萌芽枝がみられるに過ぎず,植被率は1%である.草本層には高さ0.1mほどの小型の草本が広がり,部分的に草丈の高い草本が0.3m程度の高さで生育している.部分的に植被がまばらな場所もあるため,植被率は65%である.小型の草本種としては,ツルシロカネソウ,セントウソウ,ヒメウワバミソウ,ヤマトグサ,コガネネコノメソウ,キクムグラなどが多い.高茎草本としては,アズマヤマアザミ,モミジガサ,ホウチャクソウなどが出現したが,モミジガサやホウチャクソウは小型化が著しく,開花個体はみられなかった.草丈が0.1mを超えているのは,主にシカ不嗜好性植物であるフタリシズカ,ヒロハノハネガヤ,シロヨメナ,バイケイソウ,ヤマシャクヤクなどであった.この調査区の斜面上側には,バイケイソウが大群落を形成しているが,調査区内はまだ数個体しか侵入していない.人里の半陰地に多いアシボソ,ハナタデ,ムラサキケマン,ミズヒキ,ヘビイチゴなども生育していた.


「まなびの森」の20年の活動をまとめたPDFです
![]()