








































モニタリング活動
植生調査 2014年度報告書
1.はじめに
富士山南麓では1996年(平成8年)9月の台風17号により大規模な風倒被害が発生し,壮齢ヒノキ林を中心に約90,000m3が被害を受けた.富士山「まなびの森」事業計画では,この風倒被害跡地に広葉樹を中心とした在来樹種を植栽することによって,早期に自然林を復元することを目指している.
本調査は,「まなびの森」区域内での台風による風倒跡地の森林の回復および変化について,次の3つの視点からモニタリングをおこない,自然林の早期再生のための管理・施業方法に関する情報を得ようとするものである.
1)風倒被害跡地の群状植栽地の森林回復過程について,植栽木の生育状況,群落構造の発達,林床を含めた種組成の変化を中心にモニタリングをおこなう.同時に区域内のブナ自然林,ヒノキおよびウラジロモミ人工林についても同じ項目をモニタリングし,これらとの比較から群状植栽地の回復状態を評価する.
2)風倒跡地に樹木を植栽しない場合の植生変化のモニタリングをおこない,天然更新の可能性を探索する.また調査地周辺にはニホンジカが生息するため,シカの採食圧が森林の回復にどの程度影響するかを明らかにする.
3)隣接ヒノキ林の風倒にともなうブナ自然林の質的変化をモニタリングし,風倒が周辺域に及ぼす二次的影響について明らかにする.
「まなびの森」区域内では,シカの採食圧の増加にともなって,2009年頃からスズタケの広範囲にわたる枯死がみられるとともに,バイケイソウなどの不嗜好植物の繁茂をはじめとした植生の変化が顕著になってきた.林床植生の退行にともなって,大雨時の土壌の流出も顕著になってきている.そこで,目的2)に関連して,2013年3月にブナ自然林の一部に防鹿柵を設置し,柵の内外に新たな固定調査区を設けて,柵の設置による林床植生保護の効果を検証するための調査を開始した.今年度は,この調査区の設置2年目の状況について追跡調査を行った.
2.調査区の概要
2013年3月に,ブナ自然林の固定調査区から約50m東側の緩斜面に,およそ13m×13mの防鹿柵を設置した(写真1).ネットはステン線入りナイロン製,目開き径5cmで,高さ2mで支柱間隔は約3mである.この内部に10m×10mの固定調査区を設置した.また,防鹿柵の効果を検証するための比較対象として,柵に近接する場所に同じサイズの対照区を設置した.

写真1.ブナ林内に設置された防鹿柵.
3.調査方法
2013年度の調査時期と合わせて春季(6月8日)と秋季(10月2日)の2回,防鹿柵内外の調査区(写真2)で植物社会学的な方法による植生調査をおこなった.これは,出現するすべての植物をリストアップして,それぞれの種の優占度および群度を判定するものである.優占度・群度はBraun-Blanquet(1964)の判定基準にもとづき,以下のように判定した.草本種については開花の有無も記録した.
優占度(種の個体数と被度を表す)
| r | : | 単独で生育 |
| + | : | まばらに生育し被度はごく小さい |
| 1 | : | 個体数は多いが被度は小さい(1/10以下) |
| 2 | : | 非常に個体数が多い,または被度が1/10~1/4 |
| 3 | : | 被度が1/4~1/2 |
| 4 | : | 被度が1/2~3/4 |
| 5 | : | 被度が3/4以上 |
群度(種の群がりぐあいを表す)
| 1 | : | 単独で生育 |
| 2 | : | 小群状または束状に生育 |
| 3 | : | 斑状またはクッション状に生育 |
| 4 | : | 大きな斑状,または穴のあいたカーペット状に生育 |
| 5 | : | 一面に生育 |
また,個々の草本種のサイズや開花率に対するシカの採食の影響を知るため,調査区内の代表的な草本7種(フタリシズカ,アズマヤマアザミ,モミジガサ,ホウチャクソウ,カノツメソウ,バイケイソウ,シロヨメナ)について,高さと開花の有無,食痕の有無を記録し,柵の内外で比較した.これらのうち,フタリシズカについては,茎の頂部につく花序の有無が高さに影響するため,花序を除いた茎の最上部までの高さを測定した.ホウチャクソウについては,茎が湾曲して斜上するため,茎を伸ばした長さを測定して「高さ」とした.その他の種については,花序までを含めた高さを測定した.バイケイソウは10月には地上部が枯死していたため,春季のみ測定を行った.ホウチャクソウとシロヨメナについては,秋季のみ測定を行った.
なお,測定した植物のうち,フタリシズカ,バイケイソウ,シロヨメナの3種は,富士山周辺では明らかにシカが採食を避けている不嗜好植物である.
2013年6月

2013年10月

2014年10月

写真2.
防鹿柵内(左)と柵外(右)の調査実施時の林床植生.
2014年6月は悪天候のため撮影できなかった.
4.調査結果
4.1 防鹿柵内外での種組成と開花植物の違い
表1に2013年からの計4回の調査における出現種とその優占度を示す.
柵内外の調査区は,いずれも高木層はブナの樹冠によって覆われ,シカの採食により高さ2m以下の低木層第2層がほとんど消失していること,林床にはシカの不嗜性好植物を除くと小型の植物が広がっていることなど,構造的にも組成的もほぼ同質である.ただし,低木第1層(高さ2~6m)の植被率は,柵内で50%,柵外で25%と差があった.柵外の調査区では,低木の枯死幹が多数みられ,低木層への採食の影響がより強く現れていた.
柵内外の調査区とも, 2013年と2014年で草本層の植被率や高さなどに見かけ上の大きな変化はなかったが,種組成レベルでは次のような変化が認められた.
柵内の調査区(FN)では,2013年には低木層には出現しなかったコクサギが出現した.これは,枯死寸前であった個体が新たな枝を再生したためである.草本層で2013年には出現しなかったが2014年に新たに出現した種は,イヌワラビ,サンショウ,ツルマサキ,ヤマウグイスカグラなどの10種であった.一方,2014年に確認できなかった種は,ミズ,ミツバコンロンソウ,エゴノキなどの6種であった.
柵外の調査区(FC)では,柵内とは逆に,2013年に低木層にわずかにみられたコクサギが,2014年にはみられなくなった.草本層で2014年に新たに出現した種は,タニギキョウ,ギンラン,ブナなど10種であった.2014年に確認できなかった種は,ミツバコンロンソウ,アキノキリンソウ,ウバユリなど8種であった.
草本層の構成種の開花状況についてみると,27種の植物は柵内でも柵外でも開花(または結実)が観察された(表1).このうち多くは,草丈が5cm未満でも開花することができる小型の植物か,茎が地表を匍匐する植物であり,草丈が少なくとも10cm以上にならないと開花しない高茎草本は,ホソバテンナンショウ,フタリシズカ,アズマヤマアザミ,イヌヤマハッカ,シロヨメナ,ダイコンソウ,ヒロハノハネガヤ,ミズヒキの8種のみであった.このうち,ホソバテンナンショウ,フタリシズカ,シロヨメナ,ヒロハノハネガヤの4種は,まなびの森付近ではシカの採食痕跡がほとんどみられない,明らかな不嗜好植物である.
柵内でのみ開花が観察された植物は12種あり,このうちエイザンスミレ,オオイトスゲ,タニギキョウ,ミヤマヤブタバコ,モミジガサ,ギンランの6種は,柵外の調査区にも出現はするが,開花はしていなかったものである.一方,柵外でのみ開花が確認された植物は,ムカゴイラクサ,ムラサキケマンなどの7種であったが,これらはすべて優占度+で量的には少なかった.
このほか,ウバユリ,シコクスミレ,バライチゴ,ホウチャクソウなどの草本種は柵内外の調査区に出現したが,どちらの調査区でも開花が確認できなかった.

4.2 防鹿柵内外での草本種のサイズと開花率の比較
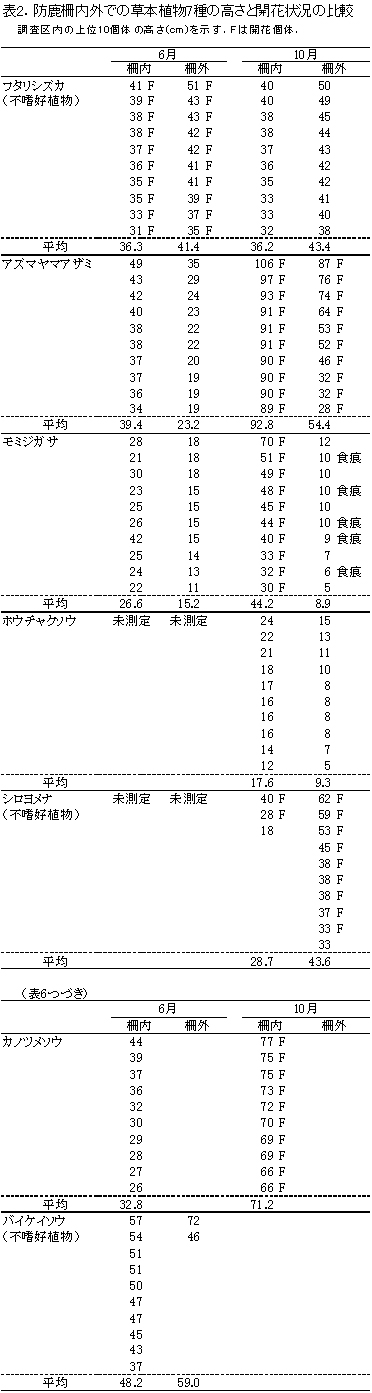
測定した7種の草本(写真3)の高さを表2に示す.このうち柵内外での比較が可能な4種の高さの平均値について,図1に示す.
フタリシズカは,春に茎頂に4枚の葉を展開し,その上に数本の花序をつける.花をつけた後はそれ以上伸長しないため,高さは6月,10月とも柵内では平均で約36cm,柵外では41~43cmであり,ほとんど変わっていなかった.柵の内外で比べると,むしろ柵外の方がやや大きな値を示していた.また,6月には計測したすべての個体が花序をつけていた.
アズマヤマアザミは,6月の時点で柵外の23.2cmに対して柵内では39.4cmと,柵内の方で草丈が高かった.10月になると柵外の54.4cmに対して柵内では92.8cmと,柵外の2倍近い高さになった.しかし,上位10個体の開花には違いがみられず,柵外の個体でもすべて開花していた.
モミジガサは,アズマヤマアザミと同様,6月の時点で柵外と柵内で高さに違いがあった.柵外の15.2cmに対して柵内では26.6cmと約10cmも大きかった.10月になると柵外ではさらに高さが低下して8.9cmとなったが,柵内では44.2cmまで成長した.柵外では,測定した個体の半数にシカの採食痕跡が認められ,夏の間に採食されていることが明らかであった.柵内の個体はすべて開花していたが,柵外では開花はみられなかった.
ホウチャクソウは,6月は未測定であったが,10月の測定値を比較すると,柵外の9.3cmに対して,柵内では17.6cmとおよそ2倍の高さがあった.
シロヨメナも6月は未測定であり,柵内の測定個体が3個体しかないため正確な比較はできないが,柵内よりも柵外の個体の方が大きい傾向があった.
カノツメソウは,柵内の調査区のみにしか出現せず,柵内の個体は6月から10月にかけて約2倍の高さに成長し,すべての個体が開花していた.
バイケイソウは,地上部が夏のうちに枯れるため,6月にしかみられなかった.柵外には2個体しかなかったが,最も大きい個体は柵外のものであった.
以上から,シカの採食による個体サイズの小型化や開花率への影響は種によって異なることがわかった.アズマヤマアザミ,モミジガサ,ホウチャクソウでは,採食圧による小型化が生じており,モミジガサでは開花率も低下していた.これに対して,不嗜好植物のフタリシズカでは,柵の内外で高さや開花率に違いがなく,シカの採食の影響を受けていないことがわかる.同じく不嗜好植物のバイケイソウやシロヨメナも,採食を受けるおそれがある柵外のほうで高さが大きな個体がみられることから,採食の影響は少ないと考えられた.
 フタリシズカ(不嗜好植物) |
 アズマヤマアザミ |
 モミジガサ |
 ホウチャクソウ |
 シロヨメナ(不嗜好植物) |
 バイケイソウ(不嗜好植物) |
写真3.高さと開花の有無を調査した草本種
5.考察
5.1 防鹿柵の有無による林床植生の違い
「まなびの森」における防鹿柵の効果を検証するための前提として,本地域の植生がすでに相当程度のシカの採食圧を受けていることを考慮しなければならない.「まなびの森」を含む西臼塚周辺のブナ林では,富士山の他の地域ではスズタケが林床に繁茂するのと異なり,草本型の林床となることが特徴である.1990年代以降のシカの増加によって高い採食圧を受け続けているため,現在の林床ではツルシロカネソウ,ヒメウワバミソウ,ヤマトグサといった小型の草本が主体となっているが,本来は,モミジガサ,カノツメソウのような高茎草本がもっと多く生育していたものと推定される.また,シカの首が届く範囲の低木層も食害により著しく衰退している.本調査でモニタリング対象としているブナ自然林の固定調査区でも,2001年の調査時から2012年の調査時までに,明らかに低木層の衰退が記録されている.シカが増加する以前の調査資料がないため,防鹿柵の設置によってどこまで植生が回復したのかを正確に評価することはできないが,上記のような高茎草本の増加や,低木層の発達,あるいは特定の不嗜好植物が増加しないこと,などが植生回復の指標として用いることができると考える.
表1に示すように,柵内でも柵外でも開花する植物は,地表面近くで花をつけられる種か,シカが好まない不嗜好植物が多かった.アズマヤマアザミやイヌヤマハッカも,完全な不嗜好植物ではないが,刺があって食べにくい,臭気を含むといった理由で,シカが食べ残す植物であると考えられる.現在の「まなびの森」の林床植生を構成しているのは主にこのような特徴をもつ植物であり,長い間のシカの採食圧を受けながらも,開花して種子を残すことができた植物であると考えられる.
一方で,柵内の調査区にしか出現しなかった種,柵内でしか開花していなかった種(表1)は,採食圧に対する抵抗性が低い植物である可能性が高い.モミジガサやミヤマヤブタバコは丈が高い高茎草本であり目立つために,柵外では開花前に花茎を食べられてしまったものと考えられる.カノツメソウやトチバニンジンのように,柵外の調査区には出現しない高茎草本は,柵の設置前にすでに消失していた可能性も考えられる.柵外にのみ出現,あるいは開花した種については,いずれも優占度が低いので,採食圧に強いため柵外に出現したのかどうかは,現段階では判断できない.
フタリシズカ,シロヨメナ,バイケイソウといった不嗜好植物は,柵内にも柵外にも生育していた.防鹿柵の近くではバイケイソウが大群生しており,今後調査地でも急速に拡大する可能性がある.こうした大型の不嗜好植物の繁茂は,植被が低下した地表面を雨による浸食から保護する効果が期待できる一方,他の草本種を排除して本来の種の多様性を低下させるおそれもある.フタリシズカやバイケイソウは,いまのところ柵外よりも柵内で優占度が高いが,今後,どちらでより増加していくのかは,モニタリングの重要なポイントとなろう.もし,柵内でもこうした不嗜好植物が増加して,他の植物を駆逐していくような現象が見られれば,この場所の植生が,柵の設置だけでは回復不能な段階に達していると判断せざるを得ないかもしれない.
5.2 草本種の形態と個体サイズ・開花率の回復との関係
柵の設置2年目までの追跡調査では,草本層や低木層の植被率や高さの増加といった明瞭な回復傾向は認められなかった.しかし,個々の植物種のレベルでは,個体サイズや開花率に柵の効果が現れ始めている(表2,図1).不嗜好植物のフタリシズカ,シロヨメナ,バイケイソウでは柵内で生長量や開花率が大きくなることはなかったが,シカが食べる植物では,柵内で個体サイズや開花率の回復がみられた.回復のしかたの違いには,それぞれの種の葉のつけ方や成長様式といった特性が関係しているようであった(図2).
モミジガサは,2013年の調査では柵内でも柵外でも開花が観察されなかった(表1)が,2014年には柵内で多くの個体が開花していた.このことは,この植物がシカの採食によって開花を抑制されており,採食されなければ比較的短期間で開花を再開できることを示している.柵外では6月よりも10月のほうが個体サイズが小さくなっていたことは,6月以降に丈の高い個体が採食され,丈の低い個体のみが残っていたことによると考えられる.根元まで食べられずに食痕が確認できる状態で生存していた個体が存在することからも,そのことが裏付けられる.モミジガサは,枝分かれしない茎を伸ばして数枚の大きな葉をつけるので,いちどシカに採食されると,光合成器官の損失が大きい.そのため,毎年採食されていると,地下茎への養分の蓄積が減少して,しだいに個体が小さくなり,開花できなくなるものと考えられる.
ホウチャクソウやカノツメソウもこれと同様であろう.ただし,ホウチャクソウは柵の設置後1年ではまだ開花個体はみられず,開花できるサイズに回復するには,モミジガサよりも時間がかかるようである.カノツメソウの場合は,採食による消耗がより大きく,前述のように柵外の調査区では消失してしまったのかもしれない.
これに対して,アズマヤマアザミは地際に広がる根生葉があることや,葉に刺があるため,刺が堅い根元近くの茎葉は食べ残されることにより,光合成器官をすべて失う危険が少ないと考えられる.また,採食された場合でも,残った腋芽が伸びて花序をつくることができる.モミジガサなどに比べると,個体サイズの小型化の程度や開花率の低下が小さいのは,こうした性質のためであると考えられた.
以上のように,林床草本の中でも,大きな葉を一斉に展開し,成長の途中で分枝しない形態をもつ高茎草本が,シカの採食の影響を受けやすい種であるといえる.そのため,シカの採食影響からの回復の程度を評価するためには,このような特徴をもつ草本種のサイズや開花率が,ひとつの指標となり得ると考えられた.また,シカの採食影響を早い段階で検知するためにも,これらの影響を受けやすい植物のサイズや開花率をモニタリングすることが有効であると考えられた.
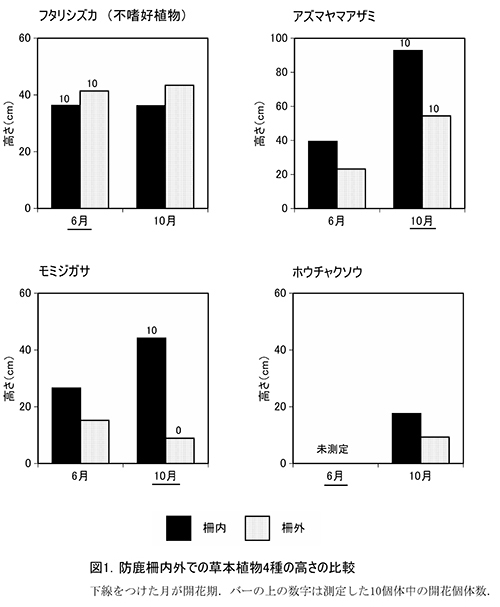
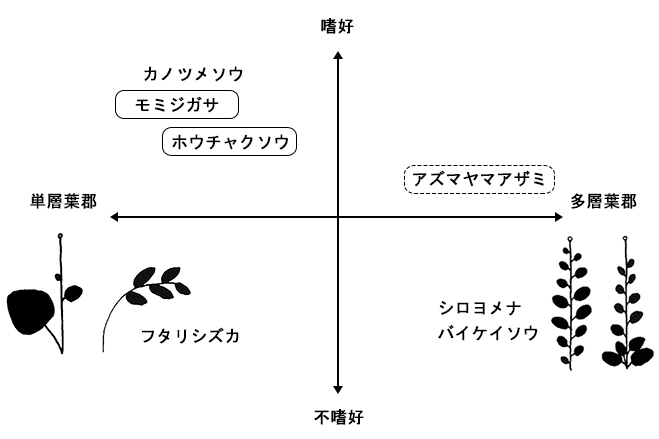
図2.シカの嗜好性,植物の形態と個体サイズ・開花への採食影響の関係

その他の種は採食による影響がみられなかった.カノツメソウは柵内にしか生育していない.


「まなびの森」の20年の活動をまとめたPDFです
![]()