








































モニタリング活動
植生調査 2015年度報告書
1. はじめに
富士山南麓では1996年(平成8年)9月の台風17号により大規模な風倒被害が発生し,壮齢ヒノキ林を中心に約90,000㎥が被害を受けた.富士山「まなびの森」事業計画では,この風倒被害跡地に広葉樹を中心とした在来樹種を植栽することによって,早期に自然林を復元することを目指している.
本調査は,「まなびの森」区域内での台風による風倒跡地の森林の回復および変化について,次の3つの視点からモニタリングをおこない,自然林の早期再生のための管理・施業方法に関する情報を得ようとするものである.
1)風倒被害跡地の群状植栽地の森林回復過程について,植栽木の生育状況,群落構造の発達,林床を含めた種組成の変化を中心にモニタリングをおこなう.同時に区域内のブナ自然林,ヒノキおよびウラジロモミ人工林についても同じ項目をモニタリングし,これらとの比較から群状植栽地の回復状態を評価する.
2)風倒跡地に樹木を植栽しない場合の植生変化のモニタリングをおこない,天然更新の可能性を探索する.また調査地周辺にはニホンジカが生息するため,シカの採食圧が森林の回復にどの程度影響するかを明らかにする.
3)隣接ヒノキ林の風倒にともなうブナ自然林の質的変化をモニタリングし,風倒が周辺域に及ぼす二次的影響について明らかにする.
2015年度は,目的2)に関連して,風倒被害発生後に樹木の植栽を行わなかった未植栽地の調査区2ヶ所について,追跡調査を行った.また,2013年3月にブナ自然林内に設置した防鹿柵による林床植生保護の効果を検証するため,防鹿柵の内外に設置した調査区2ヶ所の追跡調査を行った.固定調査区の位置を図1に,今年度モニタリングを実施した調査区の調査実施履歴を表1に示す.
2. 未植栽地の植生変化について
1. 調査方法
未植栽地の固定調査区は基線の東側,ウラジロモミ人工林に隣接した場所に位置し,10m☓10mのものを2ヶ所設置してある(図1,写真1).当初,シカの採食の有無による森林再生の状況を比較することを意図して,一方の調査区をネットで囲んだが,シカの立ち入りを防ぐことができなかったため,ネットの有無による比較は行っていない.各調査区において,2015年10月に以下の調査をおこなった.調査方法は基本的に前回までの調査と同様である.
①毎木調査
調査区内に生育している樹木について,樹高および根元直径を測定した.高木性樹種については樹高30cm以上,低木性樹種については樹高130cmの個体を調査対象とした.樹高は検測棹をもちいてcm単位で,根元直径はノギスをもちいて㎜単位で計測した.また,個体識別のためにナンバーテープをつけ,樹木の位置図を作成した.
②植生調査
植物社会学的な方法による植生調査をおこなった.これは,出現するすべての植物をリストアップして,それぞれの種に量的評価(優占度および群度)を与えるものである.優占度・群度はBraun-Blanquet(1964)の判定基準にしたがい,優占度はr,+,1~5の7段階,群度は1~5の5段階で記録した.優占度,群度の判定基準は以下の通りである.
【優占度】種の個体数と被度を表す
| r | : | 単独で生育 |
| + | : | まばらに生育し被度はごく小さい |
| 1 | : | 個体数は多いが被度は小さい(1/10以下) |
| 2 | : | 非常に個体数が多い,または被度が1/10~1/4 |
| 3 | : | 被度が1/4~1/2 |
| 4 | : | 被度が1/2~3/4 |
| 5 | : | 被度が3/4以上 |
【群度】種の生育状態を表す
| 1 | : | 単独で生育 |
| 2 | : | 小群状または束状に生育 |
| 3 | : | 斑状またはクッション状に生育 |
| 4 | : | 大きな斑状,または穴のあいたカーペット状に生育 |
| 5 | : | 一面に生育 |

ネット外

ネット内
写真1.未植栽地の固定調査区の現況(2015年10月4日撮影)
2.調査結果
(1)未植栽地(ネット外)の調査区
①樹種構成と空間分布
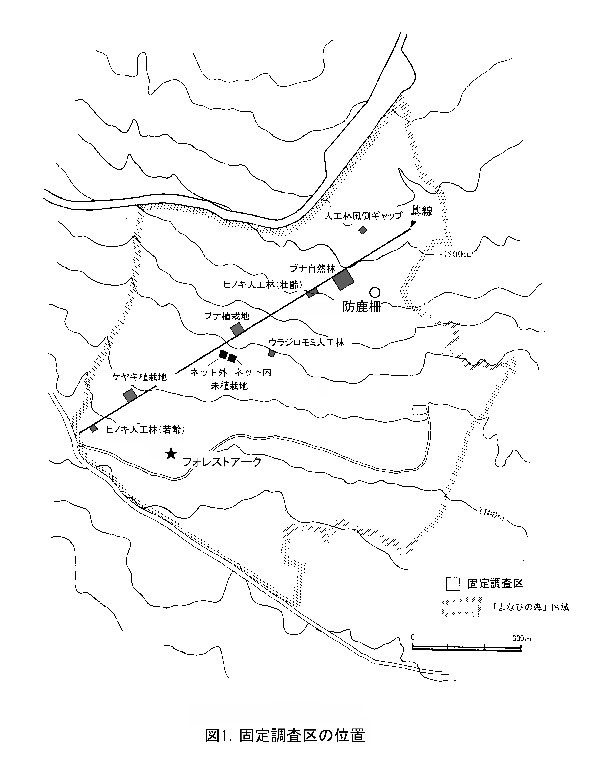
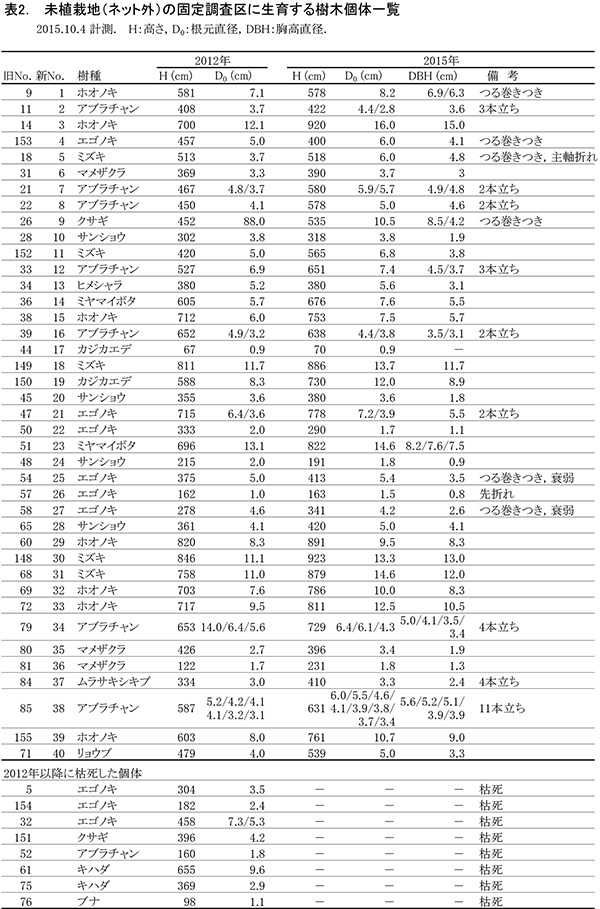
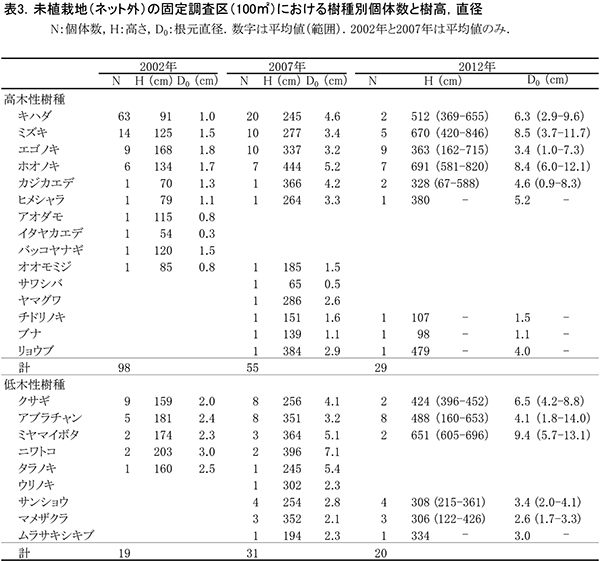
調査区内に生育している樹木の一覧を表2に,樹種別の個体数と樹高・根元直径を表3に示す.また, 2002年,2007年,2012年,および2015年の樹木位置図を図2に示す.
2002年の調査時点では,高木性樹種10種98本が生育していたが,2007年の12種55本,2012年の9種29本を経て,現在は6種22本となった(表2).樹種別ではホオノキが7本で最も多く,次いでエゴノキが6本,ミズキが5本であった.キハダは2007年には20本が生育していたが,2015年までにすべて枯死した.2012年まで生育していたチドリノキとブナも,2015年までに枯死していた.また,エゴノキも前回2012年から3本減少した.ミズキ,ホオノキ,ヒメシャラなどは増減がなかった.今回,新たに出現した高木性樹種はなかった.
低木性樹種のうち高さ130cm以上の個体は6種18本で,前回の6種20本と大きな変化はなかった(表2).アブラチャンが7本で最も多く,次いでサンショウが4本,マメザクラが3本であった.2007年に8本あったクサギは多くが枯死し,残っているのは1本のみとなった.今回,新たに出現した低木性樹種はなかった.
樹木の分布をみると,根元直径10cmを超えるホオノキ,ミズキ,エゴノキなどの高木性樹種がほぼ均等に分布していた(図2).調査区の中央付近では株立ちしたアブラチャンなどの低木性樹種の密度が高かった.
②樹木の成長と階層構造
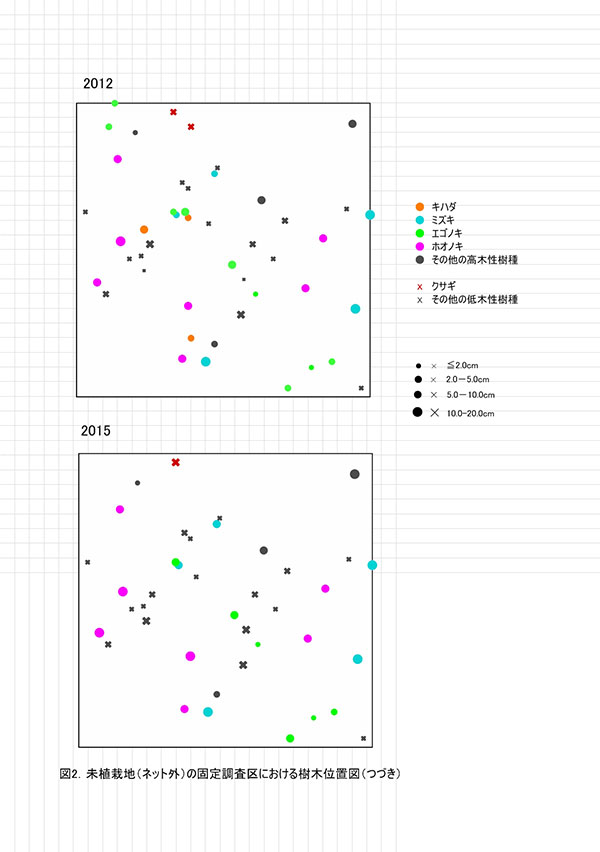
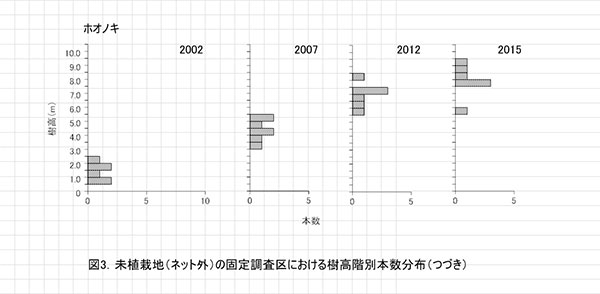
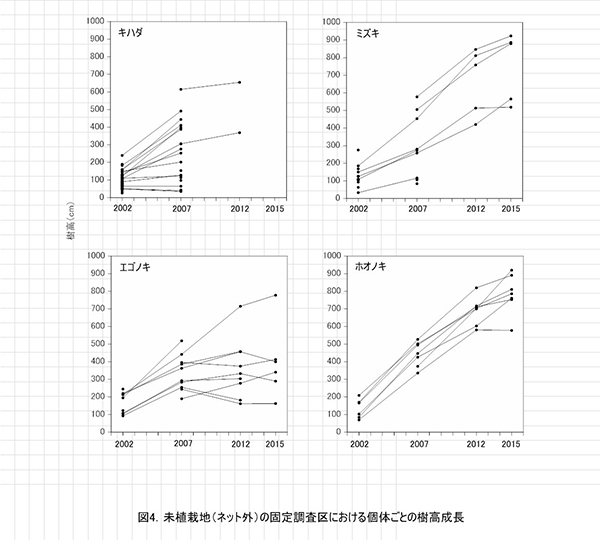
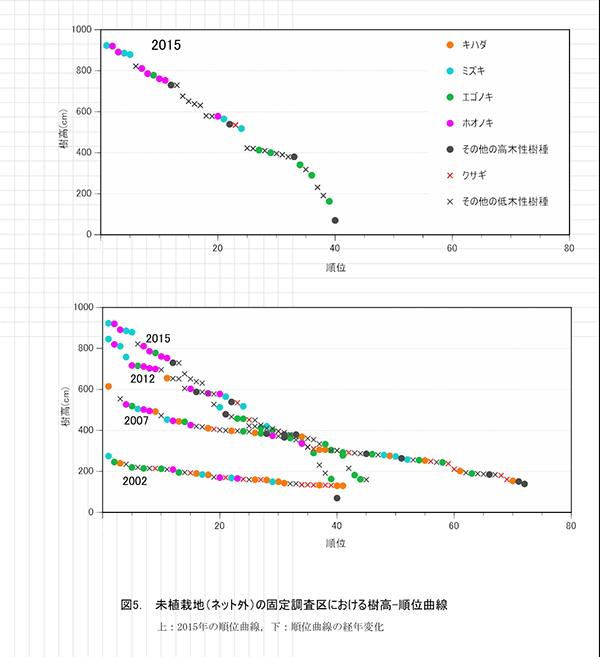
調査区内に生育する高木性樹種のうち,キハダ,ミズキ,エゴノキ,ホオノキの樹高階別本数分布の経年変化を図3に示す.図4には4種の個体別の樹高変化を示した.さらに図5には,1.3m以上の全個体の樹高を大きい順に並べた樹高順位曲線を示した.
樹高階別本数分布の経年変化をみると,ミズキでは2012年に4.5m以上であった個体すべてが生存し,その後も伸長していることがわかる.ホオノキも同様に,2012年に5.5m以上であったものが1個体を除いて,一定の割合で伸長していた(図4).一方,エゴノキでは多くの個体で伸長成長が樹高5m未満で停滞し,2012年と樹高階別の本数分布があまりかわらなかった(図3,4).これは主に,つる植物の巻きつきによって梢端が枯死したためである(表2).キハダは2012年に樹高6.5mを超えていた個体でも枯死していた.
樹高順位曲線(図5)をみると,樹高9m前後のミズキ3本とホオノキ2本が突出しており,その後に樹高7~8mのホオノキ,エゴノキなどがつづいていた.高木性樹種だけをみると,樹高7m以上のグループと6m未満のグループの間に1mほどの差があり,この間を低木性樹種のアブラチャンなどが占めている.このことから,現時点で樹高7mを超えている個体が,初期の林冠を構成するものと予想される.エゴノキは樹高4m程度で,つる植物による被陰に遭い,林冠をめぐる競争に加われなかったとみることができる.
③種組成
表4に調査区の植生調査票を示す.低木層の高さが6m近くに達し,低木の中でも下層に被圧されるものがでてきたため,低木層を二つに分け,下層を第二低木層とした.亜高木層に6種,第一低木層に15種,第二低木層に3種,草本層に67種,重複を除くと計80種の植物が記録された.
亜高木層の植被率は前回の40%から60%に増加し,ホオノキの優占度が最も高かった.そのほか,ミズキ,エゴノキ,カジカエデ,ミヤマイボタが混生していた.第一低木層の植被率は前回の60%からやや減少して50%であった.アブラチャンが最も高い優占度を占め,エゴノキ,ホオノキ,ミズキ,ヒメシャラなどの高木性樹種のほかに,クサギ,サンショウ,ムラサキシキブなどの低木性樹種が生育していた.また,つる植物のスイカズラやクマヤナギが低木層の樹冠上に広がっていた.
草本層の植被率は85%で,林冠の発達にともなって,前回の95%よりも減少した.最も優占度が高いのは,シカの不嗜好植物であるシロヨメナで,優占度4で生育していた.同じくシカの不嗜好植物であるアシボソも優占度2を維持していた.草本層では,前回の調査時に記録された種のうち17種が消失し,かわって13種が新たに出現した.消失した種には,ヨモギ,ヒメジョオン,ナギナタコウジュといった路傍雑草や,アキノウナギツカミ,キツリフネ,タニソバといった土壌攪乱のある湿地に生育する種が含まれていた.一方,新出した種には,ミヤマタニソバ,ヤマクワガタ,オタカラコウなどブナ林の構成種が含まれていた.したがって,樹木の成長にともなって,林床の構成種も徐々に森林に近づいていると考えられる.
(2)未植栽地(ネット内)の調査区
①樹種構成と空間分布
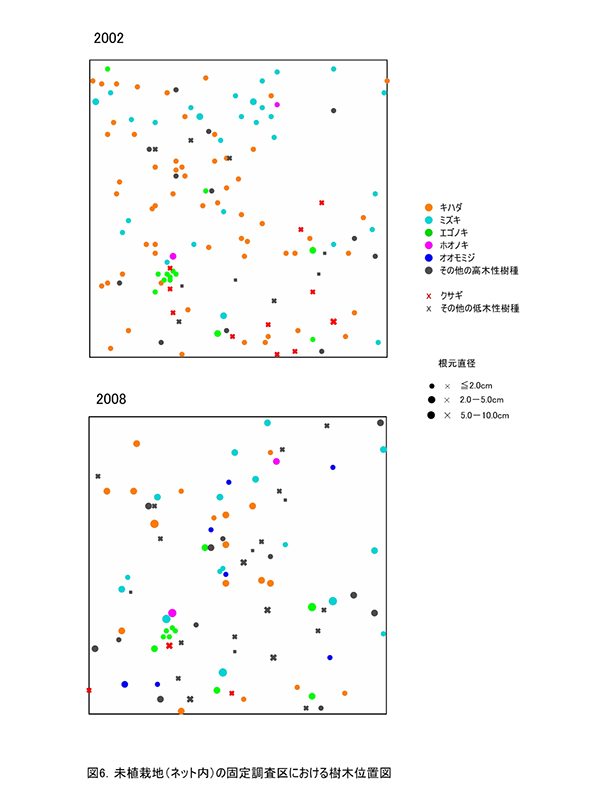
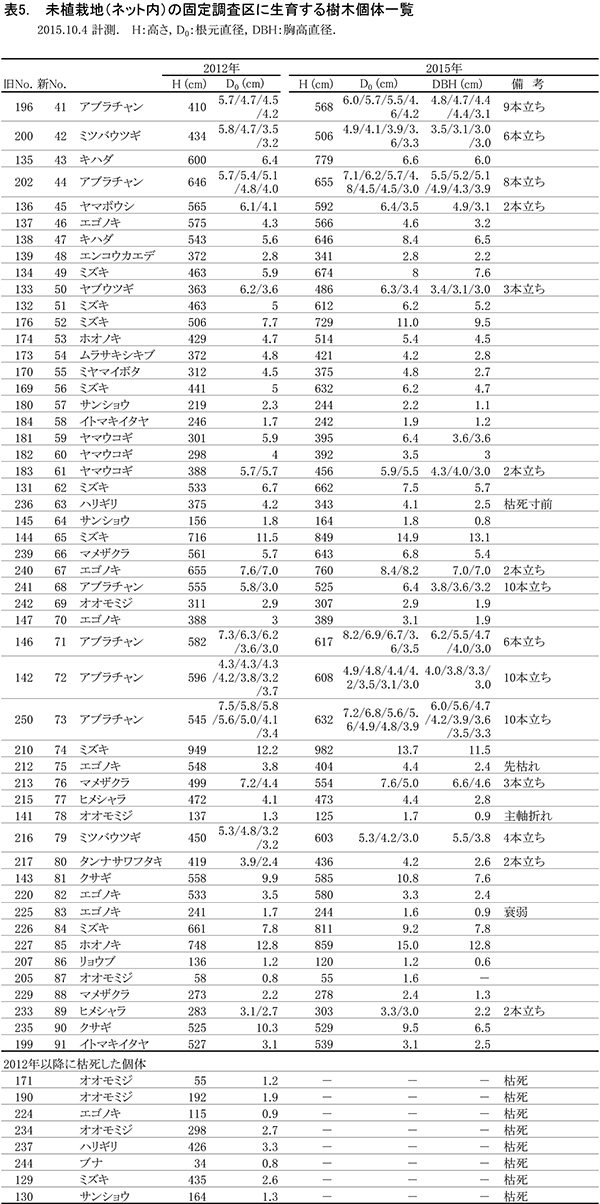
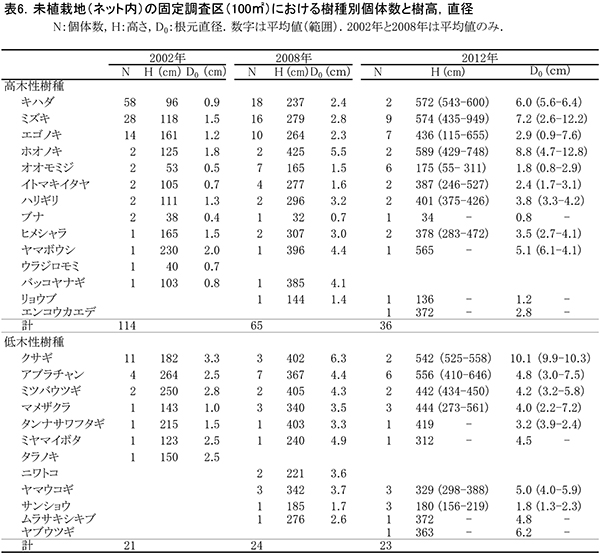
現在,調査区内に生育している樹木の一覧を表5に,樹種別の個体数と樹高・根元直径を表6に示す.また,2002年,2008年,2012年および2015年の樹木位置図を図6に示す.
2002年には高木性樹種12種114本が生育していたが,2008年の12種65本,2012年の12種36本を経て,2015年には11種29本となった(表6).最も個体数が多かったのはミズキの8本で,次いでエゴノキが6本であった.2012年に6本あったオオモミジが3本に半減していた.2002年以降,1本だけ生育していたブナは,ほとんど樹高が伸びないまま消失した.また,ハリギリは1本が残存しているが,衰弱が著しく枯死寸前であった.新たに出現した高木性樹種はなかった.
低木性樹種のうち高さ130cm以上の個体は10種22本で,2012年から1本減少しただけだった(表6).もっとも多いのはアブラチャンの6本で,ついでマメザクラとヤマウコギの3本であった.今回,新たに出現した低木性樹種はなかった.
樹木の分布は,調査区の一部で樹木を欠く部分がみられる(図6).これは,ススキが優占していた部分である.高木性樹種の分布は2012年とほとんど変わっていないが,調査区の中央部や左下部分の樹木が混み合っていた部分で,いくつかの個体が消失していることがわかる.
②樹木の成長と階層構造
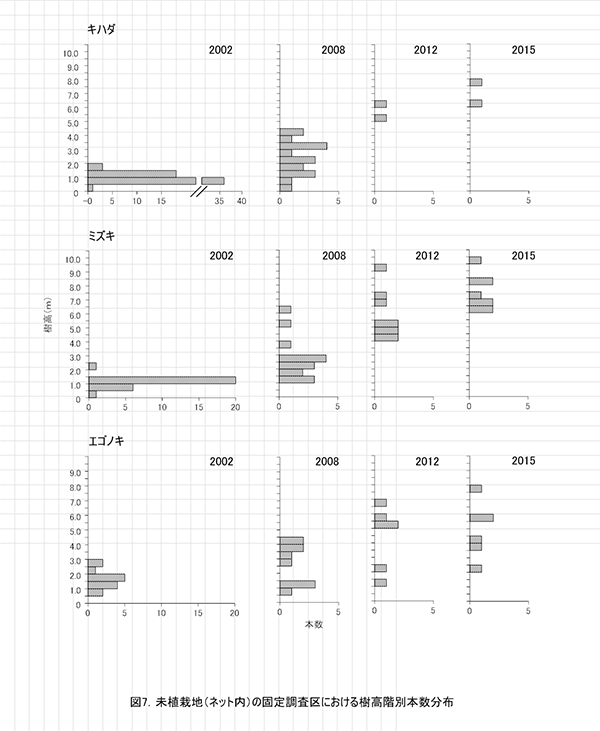
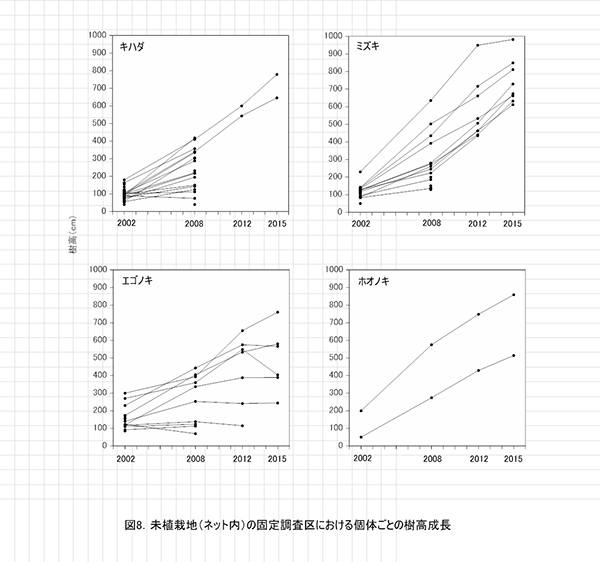
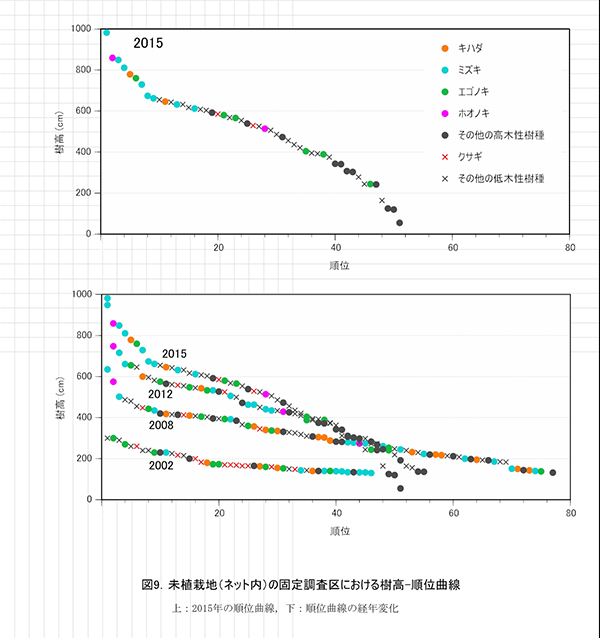
調査区内に生育する高木性樹種のうち,キハダ,ミズキ,エゴノキの樹高階別本数分布の変化を図7に示す.図8にはホオノキも含めた4種の個体別の樹高変化を示す.さらに図9は,1.3m以上の全個体の樹高を大きい順に並べた樹高順位曲線を示す.
樹高階別本数分布の経年変化(図7)をみると,いずれの樹種も2002年から2012年にかけて,樹高の大きな個体が増えているとともに,樹高のばらつきが大きくなっていた.キハダやミズキでは,樹高4m未満のものはみられなくなった.
個体識別が正確にできたものについて,2002年以降の樹高を比較すると,キハダとミズキは, 2012年の時点で生存していたものは,ミズキ1本を除いてその後も順調に成長していることがわかる(図7,8).ホオノキは当初から2本のみだったが,両方とも順調に成長していた.一方,エゴノキは,2008年の時点で樹高が4mに達していなかったものは,その後の成長が頭打ちになっていた.また,2012年以降に樹高が減少に転じたものもあった(図7,8).これはネット外の調査区と同様,つる植物の巻きつきによるものである.
樹高順位曲線(図9)をみると,ミズキ1本が樹高10m近くとなって突出しており,7~9mの間にホオノキ,ミズキ,キハダ,エゴノキが位置していた.これら7個体が初期の林冠を構成すると予想される.それ以下では曲線の傾きが緩やかになり,ミズキ4個体やアブラチャンなどの低木性樹種が続いていた.エゴノキの多くは,低木性樹種と同等の樹高にとどまっていた.オオモミジ,イトマキタヤなどのカエデ類やヒメシャラは,4m未満で低木性樹種よりも下に位置しているものが多かった.ネット外の調査区にみられた高さ4~5mでの曲線の不連続は認められず,ネット外の調査区と比べると階層の分化が進んでいない.
③種組成
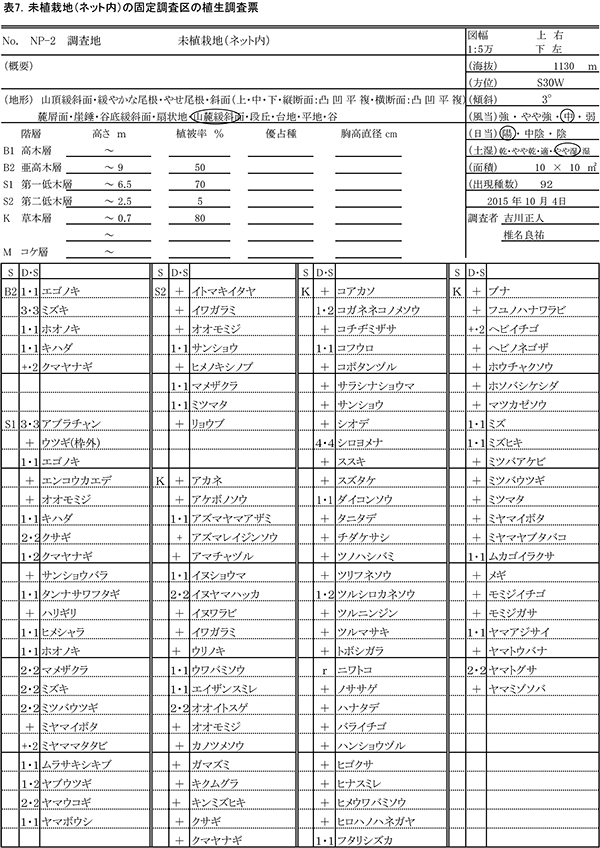
表7に調査区の植生調査票を示す.低木層の高さが6.5mに達し,低木の中でも下層に被圧されるものがでてきたため,低木層を二つに分け,2.5m以下を第二低木層とした.亜高木層に5種,第一低木層に22種,第二低木層に8種,草本層に72種,重複を除くと計92種の植物が記録された.
亜高木層の植被率は前回の15%から50%に増加し,林冠が形成されつつあることがわかる.ミズキの優占度が最も高く,そのほか,エゴノキ,ホオノキ,キハダが混生していた.第一低木層の植被率は前回の70%から変わらなかった.アブラチャンが最も高い優占度を占め,クサギ,マメザクラ,ミツバウツギ,ヤマウコギ,ヤブウツギなどの低木性樹種,ミズキ,エゴノキ,ヒメシャラなどの高木性樹種の稚樹が生育していた.また,つる植物のクマヤナギが低木層の樹冠上に広がっていた.第二低木層の植被率は5%とわずかで,サンショウ,ミツマタなどがみられた.
草本層の植被率は前回と同じ80%であった.最も優占度が高かったのは,シカの不嗜好植物であるシロヨメナで,優占度4で生育していた.その他,イヌヤマハッカ,オオイトスゲ,ヤマトグサが優占度2で生育していた.草本層では,前回の調査時に記録された種のうち12種が消失し,かわって15種が新たに出現した.消失した種には,エゴノキ,エンコウカエデ,サワシバなど木本の実生が多く,ヨモギやキツリフネもみられなくなった.ネット外の調査区で多かったアシボソも,ここでは消失していた.新出した種には,ヒメウワバミソウのような周辺のブナ林の構成種も含まれていたが,2008年から2012年の変化と比べると,森林生の植物の新たな加入は多くなかった.一方で,キンミズヒキ,トボシガラのような,林縁環境を指標する種も新出していた.
3. 防鹿柵内外の植生変化について
1.調査方法
2013年3月に,ブナ自然林の固定調査区から約50m東側の緩斜面に,およそ13m×13mの防鹿柵を設置した(写真2).ネットはステン線入りナイロン製,目開き径5cmで,高さ2mで支柱間隔は約3mである.この内部に10m×10mの固定調査区を設置した.また,防鹿柵の効果を検証するための比較対象として,柵に近接する場所に同じサイズの対照区を設置した.両調査区で,以下の調査を実施した.
①植生調査
過年度の調査時期と合わせて,春季(5月28日)と秋季(10月3日)の2回,防鹿柵内外の調査区(写真3)で植物社会学的な方法による植生調査をおこなった.優占度・群度の判定基準は,未植栽地の固定調査区で行った調査と同様である.植生調査の際,草本種については開花または結実の有無についても記録した.
②草高・開花率調査
個々の草本種のサイズや開花率に対するシカの採食の影響を知るため,春と秋の調査時に,調査区内の代表的な草本7種(フタリシズカ,アズマヤマアザミ,モミジガサ,ホウチャクソウ,カノツメソウ,バイケイソウ,シロヨメナ)について,高さと開花の有無,食痕の有無を記録し,柵の内外で比較した.これらのうち,フタリシズカについては,茎の頂部につく花序の有無が高さに影響するため,花序を除いた茎の最上部までの高さを測定した.ホウチャクソウについては,茎が湾曲して斜上するため,茎を伸ばした長さを測定して「高さ」とした.その他の種については,花序までを含めた高さを測定した.バイケイソウは10月には地上部が枯死していたため,春季のみ測定を行った.
なお,測定した植物のうち,フタリシズカ,バイケイソウ,シロヨメナの3種は,富士山周辺では明らかにシカが採食を避けている不嗜好植物である.

写真2.ブナ林内に設置された防鹿柵
2013年6月
2013年10月
写真3.防鹿柵内(左)と柵外(右)の調査実施時の林床植生
2014年10月
2015年5月
2015年10月
写真3.防鹿柵内(左)と柵外(右)の調査実施時の林床植生(つづき)
2014年6月は悪天候のため撮影できなかった.
2.調査結果
(1)防鹿柵内外での種組成と開花植物の違い
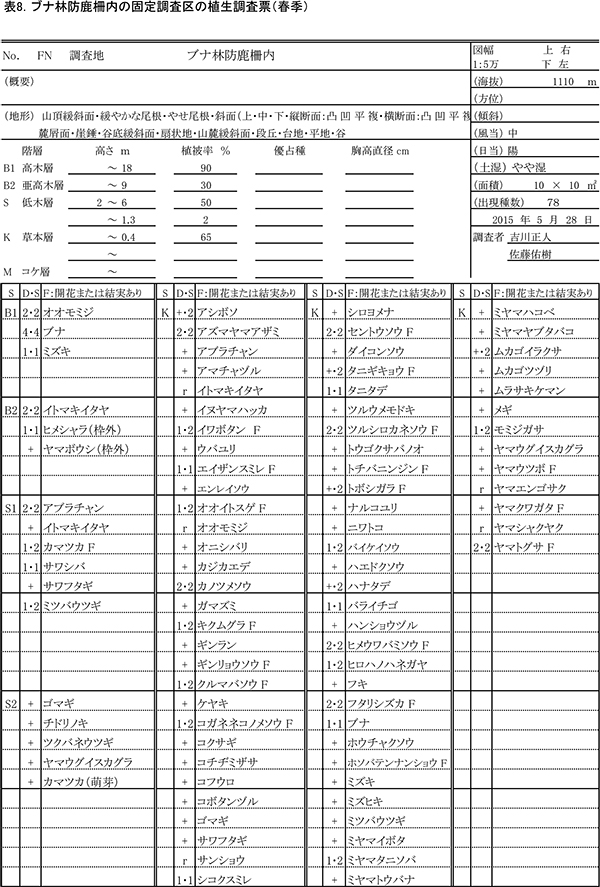
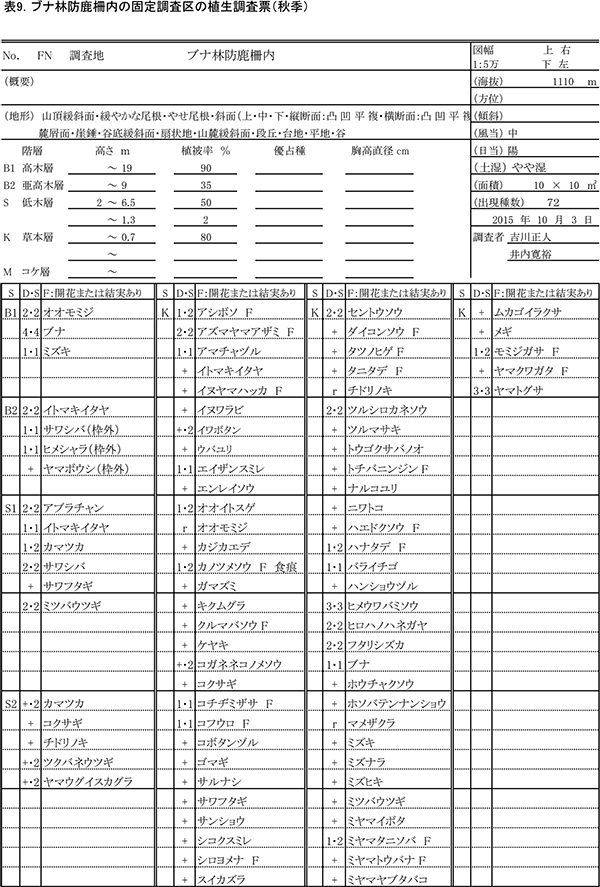
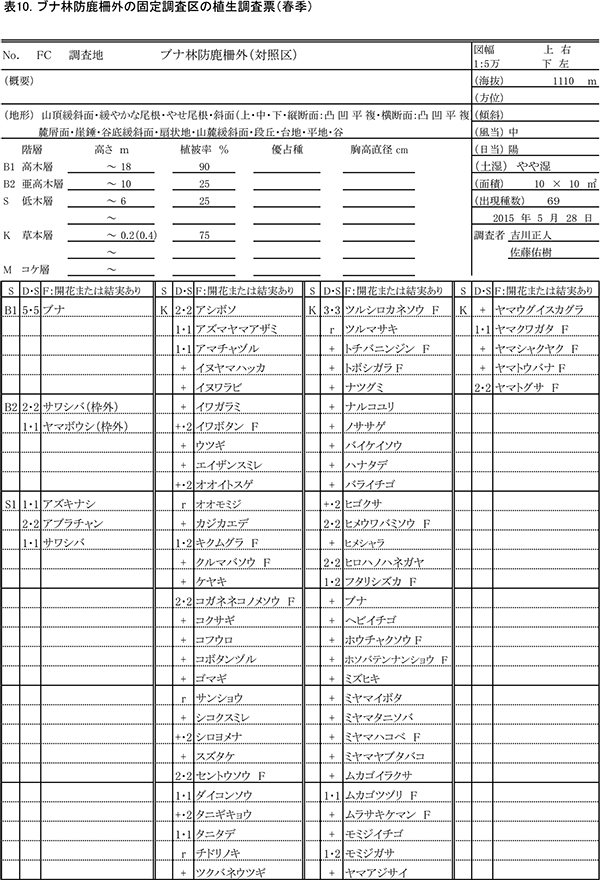
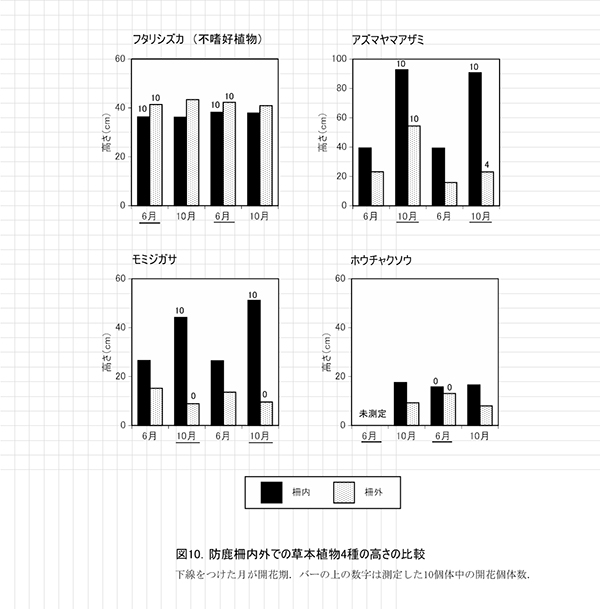
表8,9に防鹿柵内の調査区における春季と秋季の植生調査票を,表10,11に防鹿柵外の調査区の植生調査票を示す.また,表12に2013年からの計6回の調査における出現種とその優占度を示す.
柵内外の調査区は,いずれも高木層はブナの樹冠によって覆われ,シカの採食により高さ2m以下の低木第2層がほとんど消失していること,林床にはシカの不嗜性好植物を除くと小型の植物が広がっていることなど,構造的にも組成的もほぼ同質である.ただし,低木第1層(高さ2~6m)の植被率は,柵内で50%,柵外で25%と差があった.出現種数は,柵内では春季に78種,秋季に72種だったのに対し,柵外では春季に69種,秋季に62種と,10種ほど少なかった.
柵内外の調査区とも,2013年以降,草本層の植被率や高さなどに見かけ上の大きな変化はなかったが,種組成レベルでは次のような変化が認められた.
柵内の調査区(FN)で2015年に新たに記録された種は,木本の実生を除くとタツノヒゲ,エンレイソウ,ヤマトウバナ,ハンショウヅル,スイカズラの5種であった.前年に出現して2015年に出現しなかった草本種はなかった.
柵外の調査区(FC)で2015年に新たに記録された種は,木本の実生を除くとギンリョウソウ,ミヤマトウバナ,トチバニンジン,ナルコユリ,シオデの5種であった.前年に出現して2015年に出現しなかった草本種は,ギンラン,ユリワサビ,イケマの3種であった.また,低木層のツクバネウツギが消失した.柵外の調査区では,不嗜好性の一年生草本であるアシボソの優占度が,2014年以降増加していた.
草本層の構成種の開花状況についてみると,31種の植物は柵内でも柵外でも開花(または結実)が観察された(表12).このうち多くは,草丈が5cm未満でも開花することができる小型の植物か,茎が地表を匍匐する植物であり,草丈が少なくとも10cm以上にならないと開花しない高茎草本は,ホソバテンナンショウ,フタリシズカ,アズマヤマアザミ,イヌヤマハッカ,シロヨメナ,ダイコンソウ,ヒロハノハネガヤ,ミズヒキの8種のみであった.このうち,ホソバテンナンショウ,フタリシズカ,シロヨメナ,ヒロハノハネガヤの4種は,まなびの森付近ではシカの採食痕跡がほとんどみられない,明らかな不嗜好植物である.
柵内でのみ開花が観察された植物は10種あり,このうちエイザンスミレ,オオイトスゲ,タニギキョウ,モミジガサ,ギンランの5種は,柵外の調査区にも出現はするが,開花はしていなかったものである.一方,柵外でのみ開花が確認された植物は8種あったが,これらは優占度+で量的に少ないか,ムカゴツヅリやムラサキケマンのように春季にのみみられる種であった.
このほか,ウバユリ,シコクスミレ,バライチゴなどの草本種は柵内外の調査区に出現したが,どちらの調査区でも開花が確認できなかった.
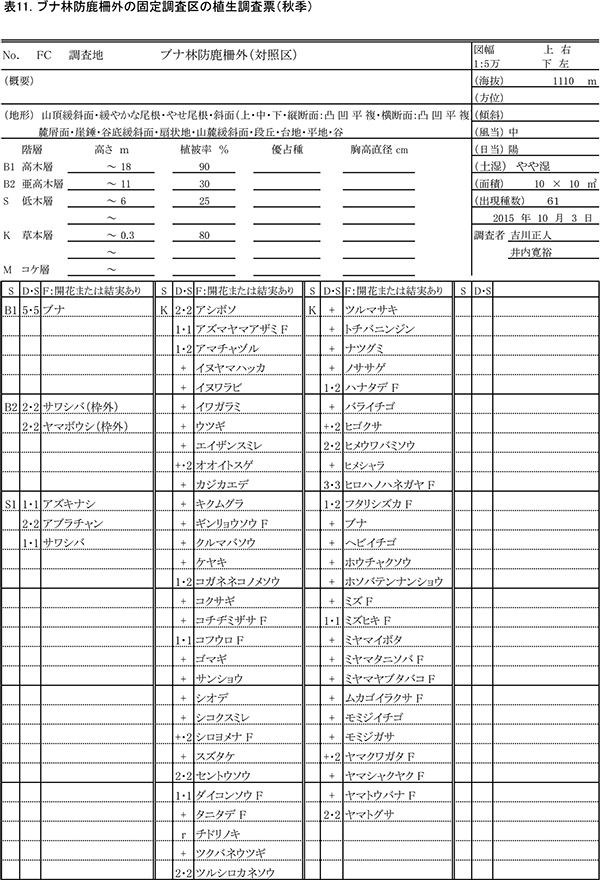

(2)防鹿柵内外での草本種のサイズと開花率の比較
測定した7種の草本の高さを,2014年の測定値とともに表13に示す.このうち柵内外での比較が可能な4種の高さの平均値の変化について図10に示す.
フタリシズカは,春に茎頂に4枚の葉を展開し,その上に数本の花序をつける.花をつけた後はそれ以上伸長しないため,高さは6月,10月とも柵内では平均で約36~38cm,柵外では約41~43cmであり,2年間でほとんど変わっていなかった.また,6月には計測したすべての個体が花序をつけていた.このことから,柵外でもシカの採食をほとんど受けていないことがわかる.
アズマヤマアザミは,春から秋にかけて茎を伸ばしながら,順次葉を展開し,葉腋に花をつけていく草本である.2014年,2015年とも,柵内では6月に40cm程度,10月に90cm程度に成長していた.しかし,柵外では6月,10月とも柵内よりも高さが低く,特に2015年には,6月から10月にかけて平均で8cmほどしか成長していなかった.また,2014年には柵外でも10個体すべてが開花していたが,2015年には開花は4個体のみであった.このことから,柵外では夏季の成長途中にシカによる採食を前年より強く受けていたと考えられる.
モミジガサは,6月頃までにほとんどの葉を展開し,秋に茎の頂部に花をつける草本である.2014年,2015年とも,6月には柵内で約27cm,柵外では13~15cmであった.10月には柵内で40cmを超えるのに対し,柵外では10cm未満に低下していた.多くの個体に食痕がみられ,夏季の間にシカに食べられていることが明らかである.モミジガサは,茎の先端を食べられると,枝を出して再生することができないため,開花することができない.柵内では10個体が開花していたのに対し,柵外では開花はみられなかった.また,柵内の個体の10月の高さは,2014年よりも7cm大きくなっており,採食されなくなったことで,より多くの光合成産物を翌年の成長に利用できるようになっていることが示唆される.
ホウチャクソウは,春季にすべての葉を展開し,6月頃までに開花する植物である.モミジガサと同様,柵外では6月よりも10月の高さが低くなっており,シカに採食されていることがわかる.柵内でもまだ高さの回復はみられず,開花個体も観察されなかった.
以上のように,アズマヤマアザミとモミジガサは,柵内で高さや開花率が明らかに大きくなることから,シカの採食によって生長量や繁殖が抑制されていたことがわかる.これらの種にとっては,柵の設置による保護効果は大きいといえる.しかし,ホウチャクソウでは,2年間では明瞭な効果は認められなかった.
なお,2015年の夏には柵内に一時的にシカが侵入しており,柵内のカノツメソウなどに食痕が認められた.そのため,柵内のカノツメソウの高さは,2014年よりも低くなっていた.また,シロヨメナについては個体数が少ないため詳しい比較は行っていないが,柵外のほうが高さが低い傾向があった.シロヨメナは不嗜好植物と考えられているが,他に食べられる植物が少ない現状では,シロヨメナも採食の影響を受けている可能性がある.


「まなびの森」の20年の活動をまとめたPDFです
![]()