








































モニタリング活動
植生調査 2016年度報告書
1. はじめに
富士山南麓では1996年(平成8年)9月の台風17号により大規模な風倒被害が発生し,壮齢ヒノキ林を中心に約90,000㎥が被害を受けた.富士山「まなびの森」事業計画では,この風倒被害跡地に広葉樹を中心とした在来樹種を植栽することによって,早期に自然林を復元することを目指している.
本調査は,「まなびの森」区域内での台風による風倒跡地の森林の回復および変化について,次の3つの視点からモニタリングをおこない,自然林の早期再生のための管理・施業方法に関する情報を得ようとするものである.
1)風倒被害跡地の群状植栽地の森林回復過程について,植栽木の生育状況,群落構造の発達,林床を含めた種組成の変化を中心にモニタリングをおこなう.同時に区域内のブナ自然林,ヒノキおよびウラジロモミ人工林についても同じ項目をモニタリングし,これらとの比較から群状植栽地の回復状態を評価する.
2)風倒跡地に樹木を植栽しない場合の植生変化のモニタリングをおこない,天然更新の可能性を探索する.また調査地周辺にはニホンジカが生息するため,シカの採食圧が森林の回復にどの程度影響するかを明らかにする.
3)隣接ヒノキ林の風倒にともなうブナ自然林の質的変化をモニタリングし,風倒が周辺域に及ぼす二次的影響について明らかにする.
今年度の調査では上記の目的1)に関して,2000年から2001年にかけて群状植栽地に設置した固定調査区2ヶ所の追跡調査をおこなった.この結果を前回までの調査結果と比較することによって,これまでの植生変化および森林の再生状況を明らかにした.また,目的2)3)に関して,2013年にブナ自然林に設置した防鹿柵内外の固定調査区の追跡調査を行い,柵設置後4年間に生じた林床植生の変化を明らかにした.
2. 群状植栽地の植生変化
1. 調査地の概要
「まなびの森」区域内の風倒被害跡地では,1998年から2002年にかけて,周辺の自然林に生育する樹種の植栽がおこなわれている.ここでは,風倒木の伐採・搬出後,一般的な地拵えはおこなわず,伐根などの残材を集積・放置してある.この放置枝条の間に,ひとつの樹種を約20m四方の群状に植栽した(群状植栽).また,基線上500-550m付近の約1haの範囲では,風倒木の搬出後,植栽をおこなわずに自然状態での森林再生を試みている.
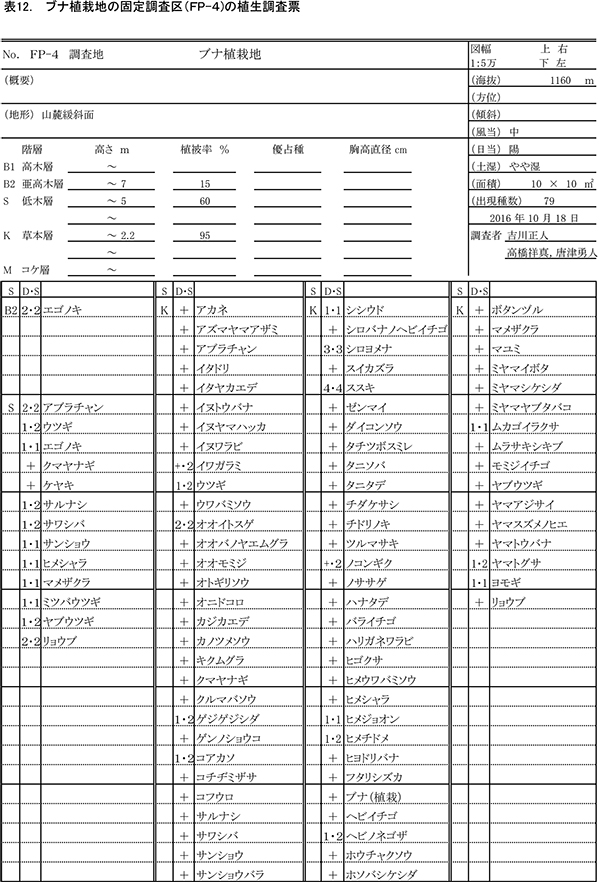
本区域内に,森林の再生状況をモニタリングするための固定調査区を9ヶ所設置し(図1),2000年以降モニタリング調査をおこなってきた.今回,調査対象としたのはケヤキ植栽地とブナ植栽地の固定調査区である.
①ケヤキ植栽地
1999年に植栽がおこなわれた場所に,翌2000年に設置した調査区であり,植栽後17年が経過している.面積は20m×20mの400㎡で,10m×10mの区画に4区分されている(ZP-1~4).調査区内には主としてケヤキが植栽されているが,コナラやイロハモミジも少数含まれている.ここでは,2000年に毎木調査,2001年に種組成の調査がおこなわれ,2003年,2007年,2010年,2013年に同じ項目が再度測定された.今回の調査は5度目の追跡調査となる.
②ブナ植栽地
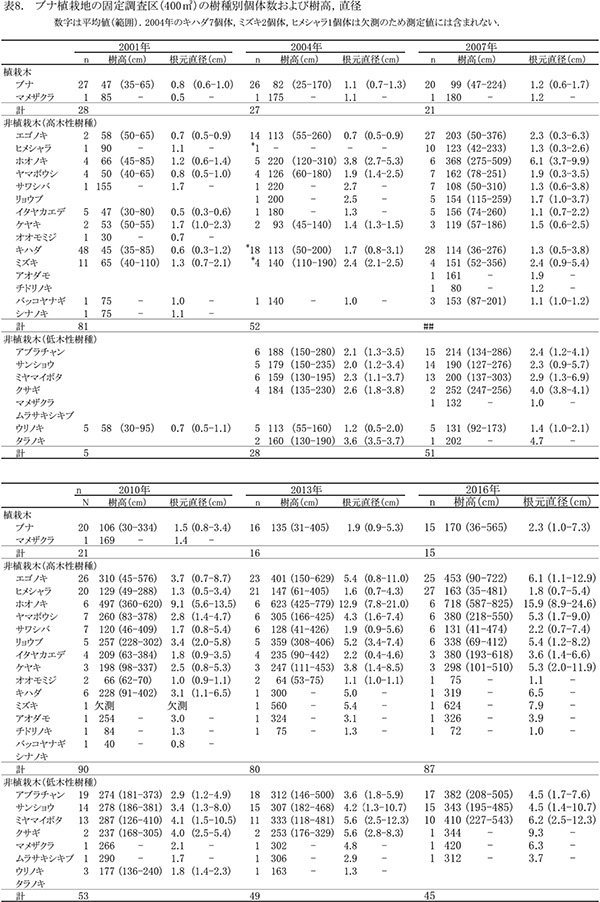
2001年に植栽がおこなわれた場所に,植栽直後の2001年に設置された調査区であり,植栽後15年が経過している.ケヤキ植栽地と同様,面積は20m×20mの400㎡で,10m×10mの区画に4区分されている(FP-1~4).調査区内には主にブナが植栽されている.ここでは,2001年に毎木調査,2002年に種組成の調査がおこなわれ,2004年,2007年,2010年,2013年に同じ項目が再度測定された.今回の調査は5度目の追跡調査となる.
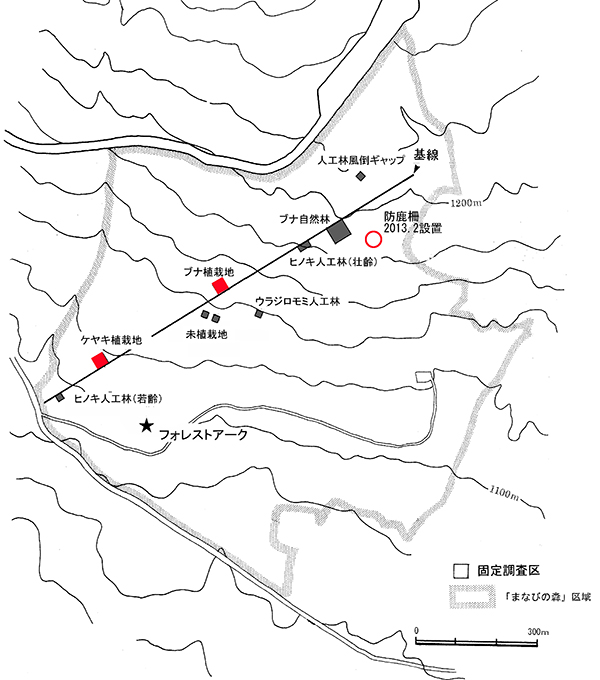
図1.固定調査区の位置
2.調査方法
①毎木調査
調査区内に生育している樹木について,植栽・非植栽にかかわらず,各個体の樹高および根元直径を測定した.高木性樹種については樹高30cm以上,低木性樹種については樹高130cmの個体を調査対象とした.樹高は検測棹をもちいてcm単位で,根元直径はノギスまたは直径巻尺を用いて㎜単位で計測した.なお,樹木の成長にともない胸高直径でのモニタリングに移行することを想定し,根元直径が5cmを超えた個体については,胸高直径(DBH,地上から1.3m高での直径)も測定した.また,個体識別のためにナンバーテープをつけ,位置図を作成した.
②植生調査
2016年10月に,10m×10mの区画を単位として,植物社会学的な方法による植生調査をおこなった.これは,出現するすべての植物をリストアップして,それぞれの種の優占度および群度を判定するものである.優占度・群度はBraun-Blanquet(1964)の判定基準にもとづき,以下のように判定した.
優占度(種の個体数と被度を表す)
| r | : | 単独で生育 |
| + | : | まばらに生育し被度はごく小さい |
| 1 | : | 個体数は多いが被度は小さい(1/10以下) |
| 2 | : | 非常に個体数が多い,または被度が1/10~1/4 |
| 3 | : | 被度が1/4~1/2 |
| 4 | : | 被度が1/2~3/4 |
| 5 | : | 被度が3/4以上 |
群度(種の群がりぐあいを表す)
| 1 | : | 単独で生育 |
| 2 | : | 小群状または束状に生育 |
| 3 | : | 斑状またはクッション状に生育 |
| 4 | : | 大きな斑状,または穴のあいたカーペット状に生育 |
| 5 | : | 一面に生育 |
3.調査結果
3-1.ケヤキ植栽地
①樹木個体数と空間分布
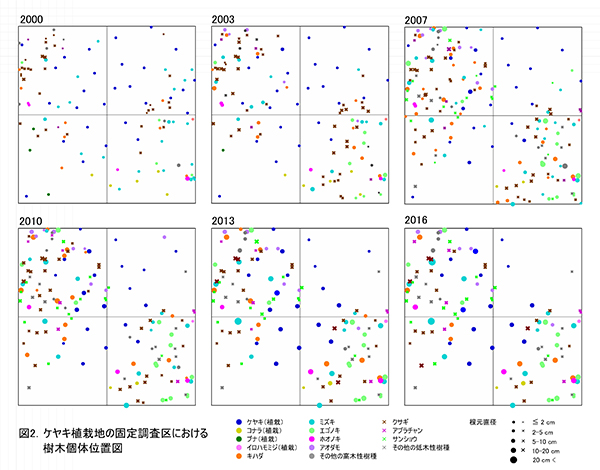
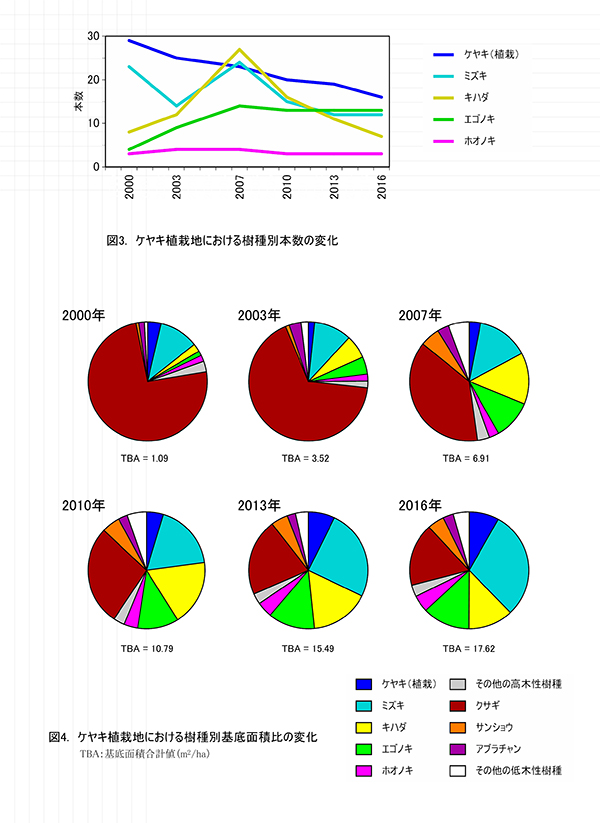
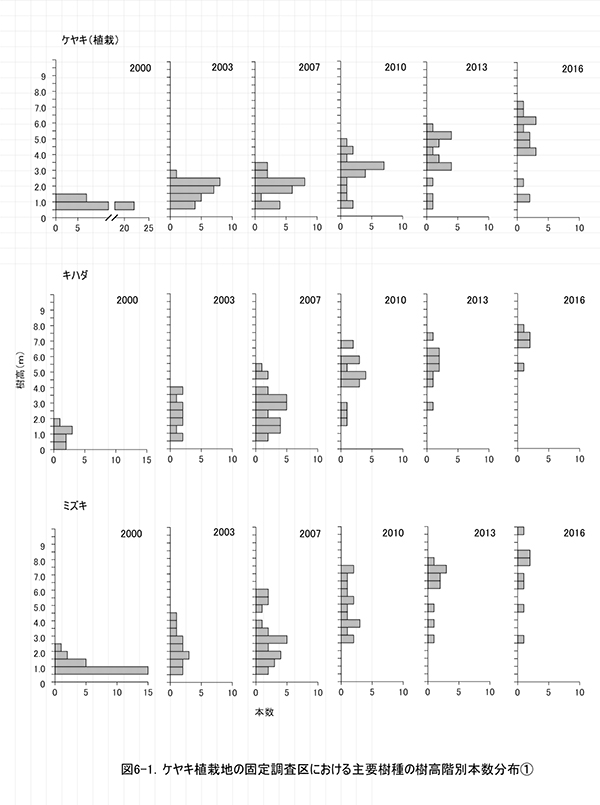
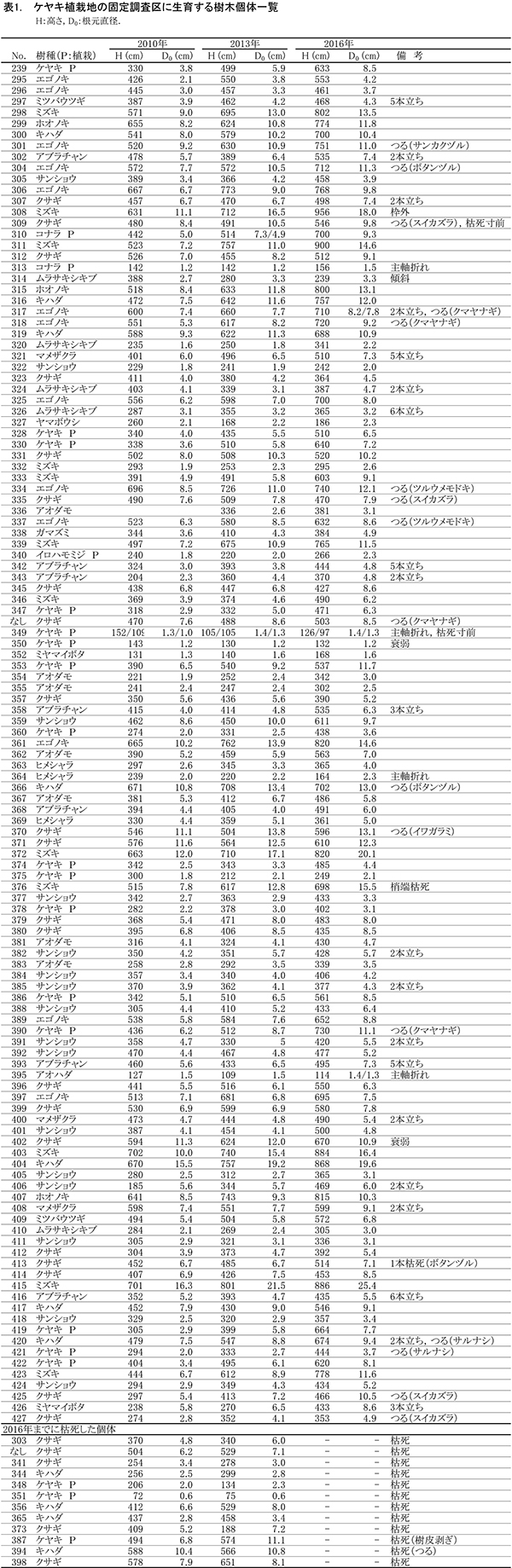
ケヤキ植栽地の固定調査区(400㎡)内に生育する樹木の一覧を表1に,樹種別の個体数と樹高・根元直径を,過去の測定結果とともに表2に示した.また,樹木の分布を図2に示す.さらに,主な樹種の本数の変化を図3に,優占度(基底面積比)の変化を図4に示す.
今回,測定対象となった樹木は123本であった.新しく記録された個体はなく,前回記録された個体のうち12本が枯死していた(表1).生存していた植栽木は,ケヤキ16本,コナラ2本,イロハモミジ1本の計19本で,前回2013年の調査以降,ケヤキ3本が枯死していた(表2).非植栽木についてみると,高木性樹種では8種47本が生育していた. 前回2013年の調査と比べると,ミズキ(12本),エゴノキ(13本),アオダモ(7本)などは本数が変化していないが,キハダだけが4本枯死して7本となった.低木性樹種では,8種57本の生育が記録された.前回2013年の調査時から5本減少したが,枯死した5本はすべてクサギであった.クサギ以外の低木は,2010年以降ほとんど本数が変化していなかった(表2).
樹木の空間分布(図2)をみると,枝条が集積された区画ZP-2とZP-3(左上と右下の区画)への偏りが一層明確になってきたことがわかる.これらの区画ではエゴノキ,ミズキ,キハダ,ホオノキの4種で,根元直径10cm以上,樹高8m以上に達するものが現れ,しだいに樹林が形成されつつある.しかし,区画ZP-4(右上の区画)では,ススキの繁茂が衰えず,植栽したケヤキがわずかに残っているほかは,天然生稚樹の成長はほとんどみられない.
主な高木性樹種の個体数の変化(図3)をみると,植栽したケヤキは徐々に減少しているが,依然としてもっとも本数が多い.2007年まで増加したミズキとキハダはその後減少し,相対的にエゴノキがケヤキに次ぐ本数となった.樹種ごとの優占度の指標として根元直径から算出した基底面積の割合の変化(図4)をみると,当初は3/4を占めていた先駆性低木のクサギが徐々に減少し,現在ではミズキ(29.8%),エゴノキ(12.9%),キハダ(12.2%)の3種が全体の半分を占めていた.ケヤキは本数は多いが,基底面積比では8.1%にとどまっていた.基底面積の合計値は,過去3年間で2.13㎡/ha増加して,17.62㎡/haとなった.これは,調査を開始した2000年の約16倍の値である.
②樹木の成長と階層構造
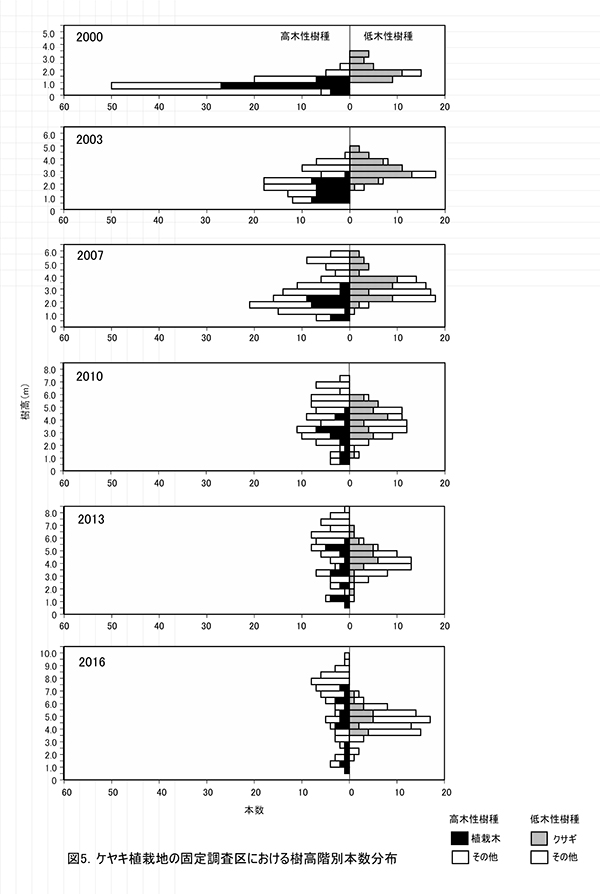
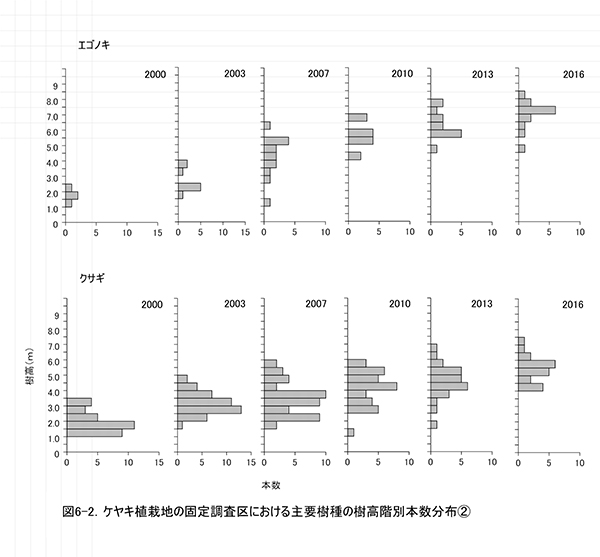
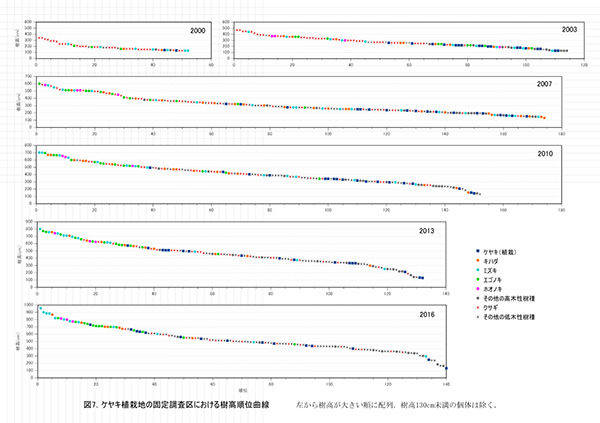
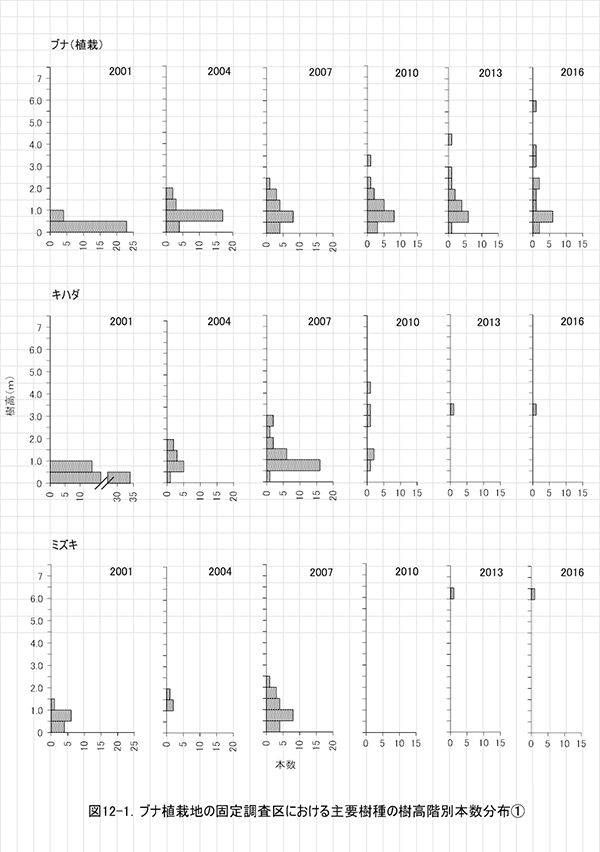
調査区内に生育する樹木の樹高階別本数分布の経年変化を図5に,そのうち個体数が多い樹種については図6に個別に示した.また,図7には1.3m以上の全個体の樹高を大きい順に並べた樹高順位曲線を示した.
全樹種の樹高階別本数分布の経年変化(図5)をみると,2000年には樹高1m未満のものが多かった高木性樹種の分布が,2003年から2007年にかけては二山形の分布になり,2010年以降のばらつきが大きい時期を経て,2016年には7.5-8.0mをピークとする一山型に近づいていることがわかる.二山形の分布が崩れたのは,高さ約3mから6m程度にクサギをはじめとする低木性樹種の階層が形成され,その下になった高木性樹種(特にミズキとキハダ)の多くが枯死したためである.現在のピークを形成しているのは,この低木性樹種の階層を抜け出した個体である.一方,低木性樹種も高さ3.5-5.5mの個体が多くなっており,高木性樹種との間ではっきりした階層構造を形成しつつある段階にあるとみられる.
樹種別(図6)にみると,ケヤキとミズキでは,個体間の高さのばらつきが大きく,まだ低木性樹種と競合する高さにとどまっている個体もある.そのため,この2種では今後も被圧により枯死する個体が生じる可能性が高い.一方,キハダとエゴノキではすでに樹高が小さい個体は枯死しており,低木性樹種との競争からは開放された個体のみが残っていると考えられる.ただし,エゴノキの場合は,ツルウメモドキ,クマヤナギ,サンカクヅルといった木本性のつる植物にからまれている個体が多く,今後はつる植物による被圧の影響が出てくるかもしれない.
樹高順位曲線(図5)では,樹高6m以上(上位40本)はほぼ高木性樹種に占められており,それ以下では低木性樹種が多いことがわかる.低木性樹種の中でも先駆的な性質が強いクサギが衰退し,他の低木性樹種も樹高限界に近づいてきたことから,この上位40位までの個体がこれから林冠を形成していくと予想される.とくにミズキの成長が良好で,上位4個体は8.5m以上に達しており,ミズキが二次林初期の優占種となる可能性が高い.
③林床植生
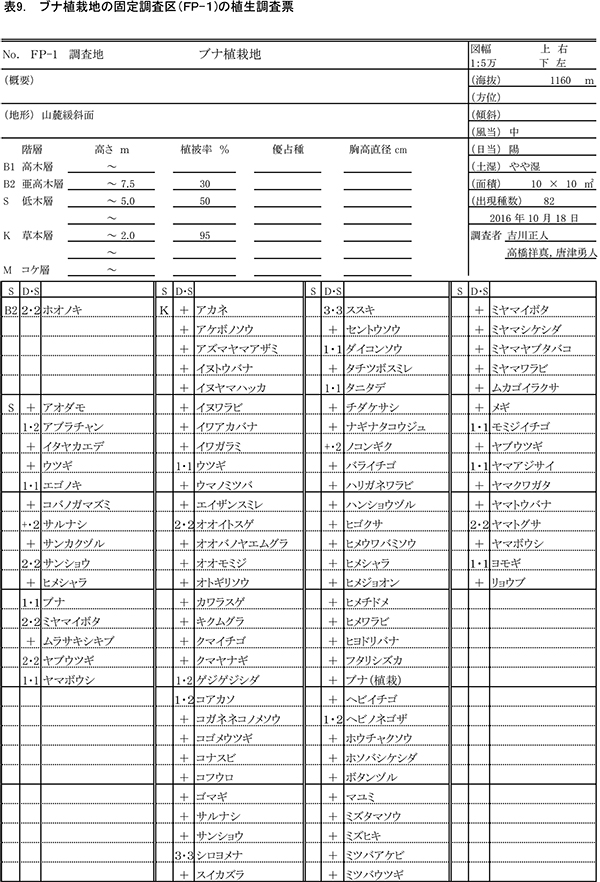
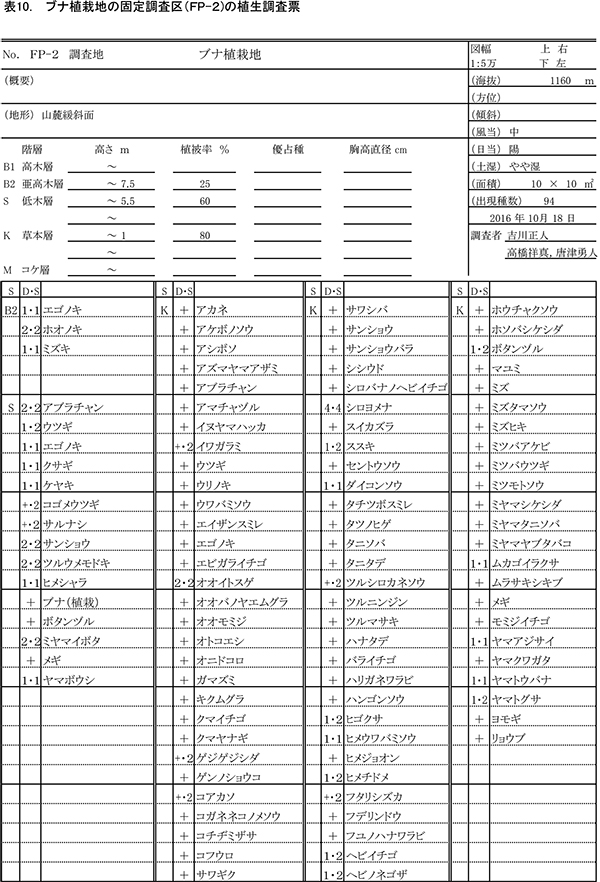
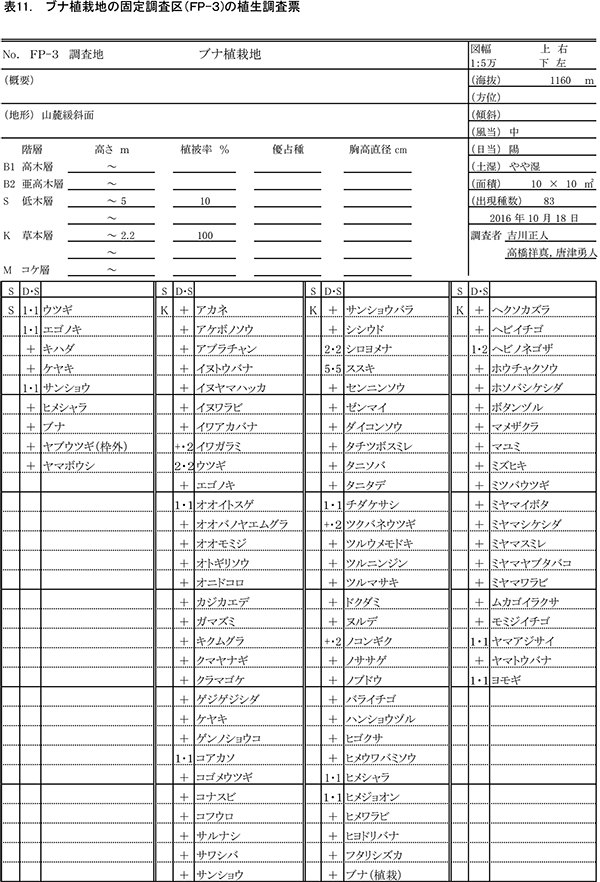
表3~6に調査区内10m×10mの4区画ごとの植生調査票を示す. 現在の調査区の植生は,おおむね6mから9mの亜高木層,2mから5.5mの低木層,0.8m以下の草本層の3階層からなる.ただし,区画ZP-4では,ススキの繁茂が著しく,草本層が2.3mに達している.区画ごとの出現種数は77種から100種であった.
亜高木層には,天然生の稚樹のうち生育のよいミズキ,キハダ,エゴノキ,ホオノキと,先駆性樹種のクサギが生育するほか,つる性木本のクマヤナギやサンカクヅルも出現した.低木層には,クサギ,ヤブウツギ,アブラチャン,サンショウ,コクサギ,ミツバウツギ,マメザクラ,ムラサキシキブなどが多く,亜高木層と同種の高木性樹種や植栽したケヤキも含まれていた.
草本層では出現種数が非常に多く,森林生の植物,草原生の植物,路傍雑草的な植物など,いろいろな種が混在している.ヒメウワバミソウ,セントウソウ,ツルシロカネソウ,シコクスミレ,コガネネコノメソウなど,周辺のブナ自然林の林床に多い植物も多数生育しており,組成的には既にかなり自然林に近づいているものと考えられる.ハリガネワラビ,ホソバシケシダなどのシダ植物は,周辺の林内にはあまりみられないが,調査区では放置枝条の周囲に生育していることが多い.シカの不嗜好性植物であるシロヨメナの優占度が,いずれの区画でも2~4と大きくなっている一方,その他の高茎草本は優占度が小さいことから,シカの採食の影響が強いと推定される.樹木の生育量が少ないZP-4ではススキが優占度5に達しており,周囲の樹木が成長して日当たりが悪くなるまでは,草原状の植生で維持されると考えられる.
3-2.ブナ植栽地
①樹木個体数と空間分布
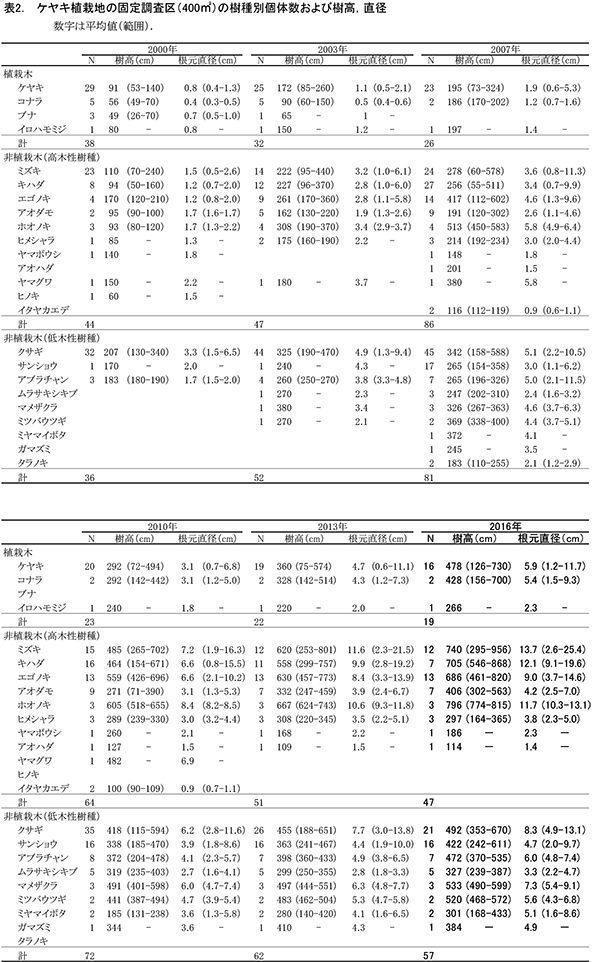
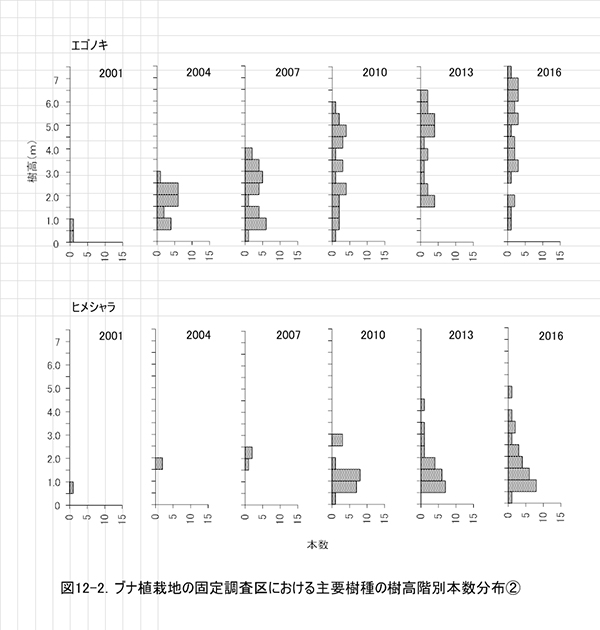
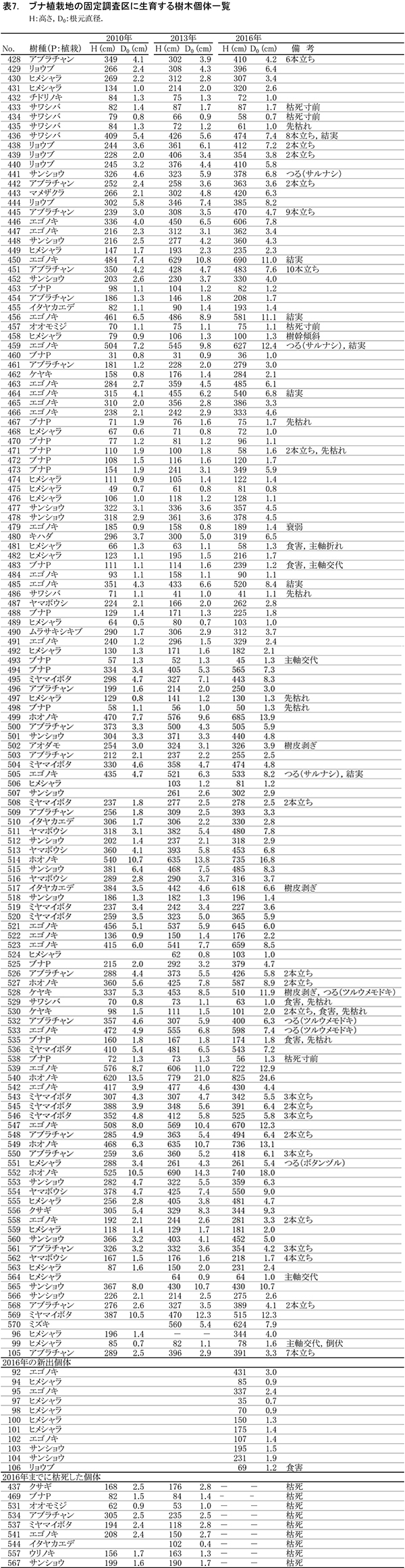
ブナ植栽地に設置した固定調査区(400㎡)内に生育する樹木の一覧を表7に,樹種別の個体数と樹高・根元直径を,過去の測定結果とともに表8に示した.また,樹木の分布を図8に示す.さらに,主な樹種の本数の変化を図9,優占度(基底面積比)の変化を図10に示す.
今回の調査で測定対象となった樹木は147個体であった.前回2013年の調査時からの新出個体は11本,枯死個体は9本で,全体で2本の増加となった(表8).植栽したブナは,前回2013年の調査時から1本が枯死して15本となった.非植栽木についてみると,高木性樹種は13種87本が生育しており,前回より7本増加した.樹種別ではヒメシャラが最も多く27本,次いでエゴノキが25本であった.多くの樹種の本数がほとんど変化しなくなる中,ヒメシャラは5本,エゴノキは3本の新出木が加入していた.また,ヒメシャラ1本(No.96)は,前回枯死したと判断されたが,根元から吹き返して再生していた.低木性樹種は6種45本で,前回2010年の49本からわずかに減少した.個体数が多い樹種は,アブラチャン17本,サンショウ15本,ミヤマイボタ10本であった.先駆的な性質が強いクサギは,1本が残るのみとなった.
樹木の空間分布(図8)をみると,樹木個体は放置枝条のある区画FP-1,FP-2,FP-4に多く,ススキが繁茂しているFP-3(図の右下の区画)ではやや少ないことがわかる.しかし,ヒメシャラとエゴノキの稚樹はFP-3の区画にもみられ,ススキが繁茂した中でも枯死せずに生育していた.
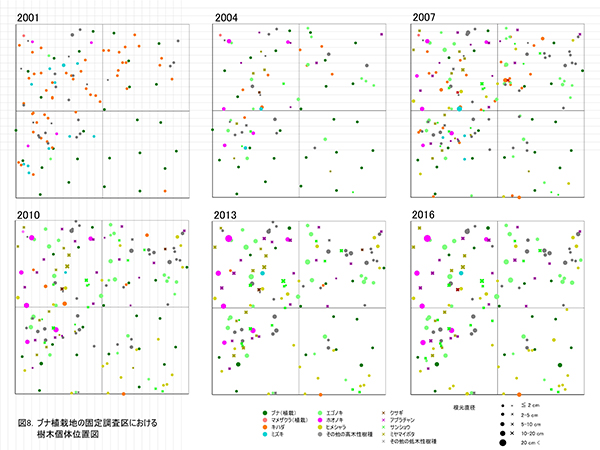
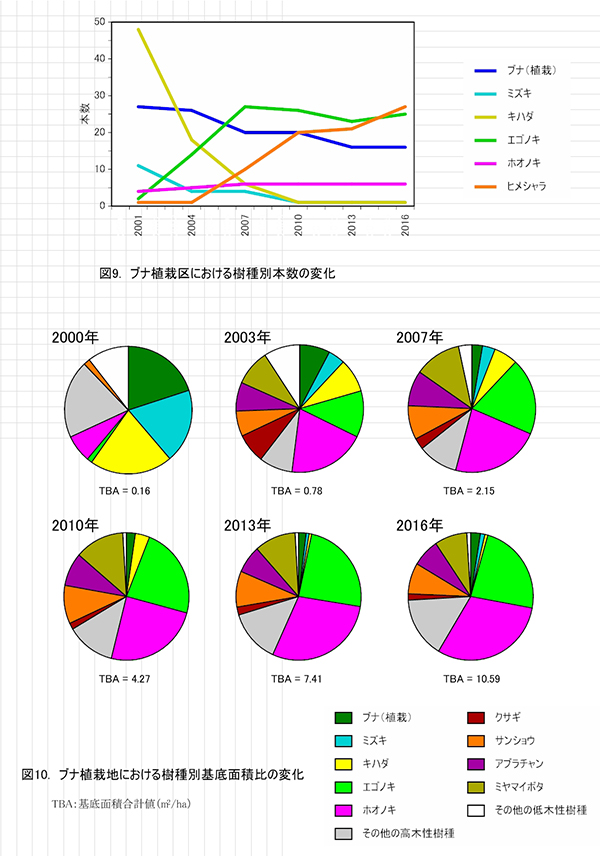
主な高木性樹種の個体数の変化(図9)をみると,ミズキやキハダが減少した一方で,ヒメシャラが徐々に増加して,2016年にはエゴノキを抜いて最も個体数が多い樹種となっており,調査開始時からは樹種構成が大きく入れ替わっている.樹種ごとの優占度の指標として根元直径から算出した基底面積の割合の変化(図10)をみると,最も大きな割合を占めているのはホオノキ(30.7%)で,次いでエゴノキ(23.5%)の割合が高く,この2種で全基底面積の半分以上を占めていた.ヒメシャラは個体数は多いが,基底面積比ではまだ全体の2.4%にすぎない.基底面積の合計値は10.59㎡/haで,ケヤキ植栽地と比べると2/3ほどであった.
②樹木の成長と階層構造
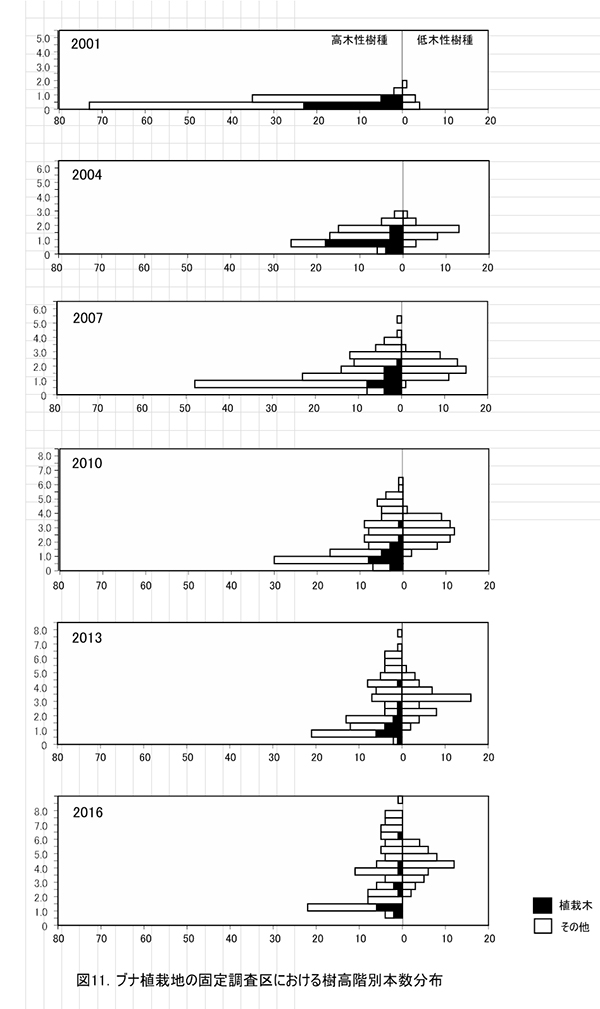
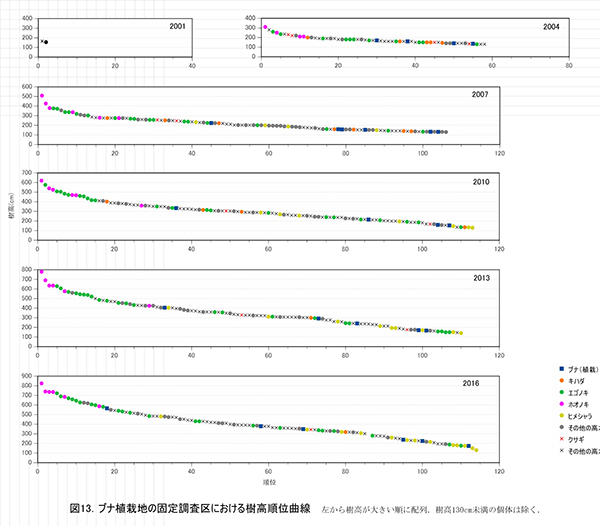
調査区内に生育する樹木の樹高階別本数分布の経年変化を図11に,そのうち主要な樹種については図12に個別に示した.また,図13には1.3m以上の全個体の樹高を大きい順に並べた樹高順位曲線を示した.
全樹種の樹高階別本数分布の変化(図11)をみると,高木性樹種の樹高にはばらつきが大きく,個体数が最も多いのは1.0~1.5mの階級である.ヒメシャラの稚樹が新たに加入していることもあり,ケヤキ植栽地と比べると樹高の小さい個体が多い.高木性樹種の成長のよい個体は6.0mを超え,低木性樹種よりも大きくなっていたが,植栽木でしたブナのほとんどは,依然として低木性樹種に高さを超えられていない.樹種別にみると,エゴノキが樹高3m以上,ヒメシャラが樹高4m未満を中心に,サイズの異なる稚樹が連続的に生育していることがわかる(図12).
樹高順位曲線の経年変化(図13)をみると,ホオノキとエゴノキが上位を占める傾向は2010年頃から変わらず,樹高5mを超える上位20本までは,ほとんどがこの2種であった.その後には,サンショウ,ミヤマイボタ,アブラチャンなどの低木性樹種が続いており,80位以下(樹高約3m)になるとヒメシャラが多くなっていた.ここでは,ホオノキとエゴノキが初期の林冠を形成していくと予想される.
③林床植生の変化
表9~12に調査区内10m×10mの4区画ごとの植生調査票を示す.現在の調査区内の植生は,一部で7.5m程度の亜高木層が形成されつつあるが,大部分は2mから5.5mの低木層と,2.2m以下の草本層に区分できる.草本層はススキが密生する階層に対応し,細かくみればその内部に1m以下の草本層第2層を認めることができるが,調査では区別しなかった.区画ごとの出現種数は79種から94種であった.
亜高木層にみられるのは,突出して成長しているホオノキ,エゴノキ,ミズキの3種のみである.低木層には,アブラチャン,サンショウ,ミヤマイボタ,ヤブウツギ,ウツギなどの低木性樹種や,エゴノキ,ヤマボウシなど高木性樹種の稚樹が生育している.ツルウメモドキ,サルナシ,サンカクヅルなどのつる植物も出現する.
草本層ではススキが優占しており,低木層が発達したFP-2(左上の区画)以外の3区画では,優占度3以上であった.また,シカの不嗜好植物であるシロヨメナも優占度2~4と多く生育していた.ケヤキ植栽地の調査区と同様,森林性のものから陽地性のものまで,さまざまな環境の種が混在していたが,ケヤキ植栽区よりも,シシウド,ヒヨドリバナ,ノコンギクなど草原生の種が多く出現した.また,ヒメウワバミソウ,ツルシロカネソウ,セントウソウなど林床生草本の出現頻度や優占度は,ケヤキ植栽区よりも小さかった.これらのことから,ブナ植栽区では,樹林への遷移の進行が,ケヤキ植栽区よりも少し前の段階にあるといえる.
3. 防鹿柵内外の植生変化
1.調査方法
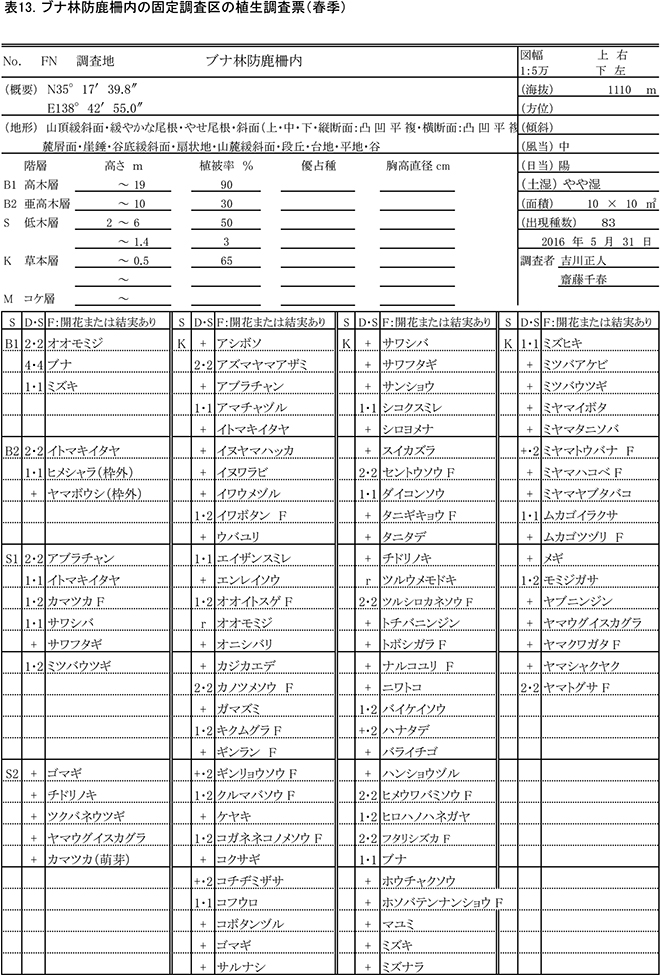
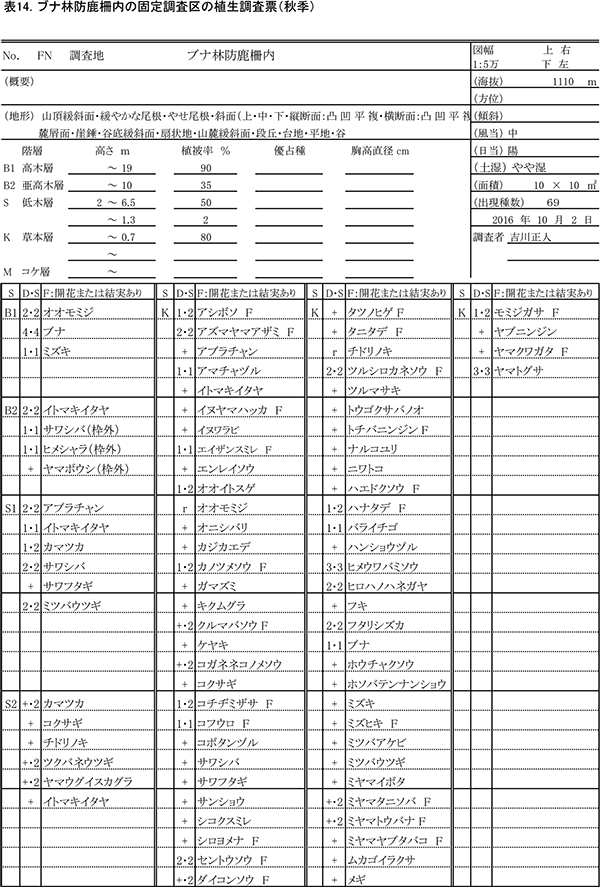
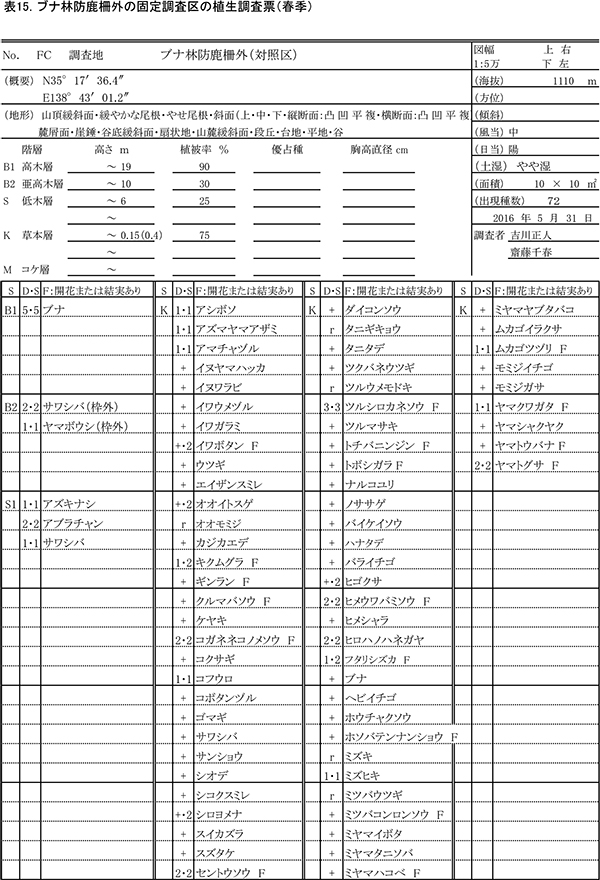
2013年3月に,ブナ自然林の固定調査区から約50m東側の緩斜面に設置された,およそ13m×13mの防鹿柵の内部に10m×10mの固定調査区を設置した.また,防鹿柵の効果を検証するための比較対象として,柵に近接する場所に同じサイズの対照区を設置した.両調査区で,以下の調査を実施した.
①植生調査
過年度の調査時期と合わせて,春季(5月31日)と秋季(10月2日)の2回,防鹿柵内外の調査区(写真1)で植物社会学的な方法による植生調査をおこなった.優占度・群度の判定基準は,植栽地の固定調査区で行った調査と同様である.植生調査の際,草本種については開花または結実の有無についても記録した.
②草高・開花率調査
個々の草本種のサイズや開花率に対するシカの採食の影響を知るため,春と秋の調査時に,調査区内の代表的な高茎草本7種(フタリシズカ,アズマヤマアザミ,モミジガサ,ホウチャクソウ,カノツメソウ,バイケイソウ,シロヨメナ)について,調査区内の上位10個体の高さと開花の有無,食痕の有無を記録し,柵の内外で比較した.これらのうち,フタリシズカについては,茎の頂部につく花序の有無が高さに影響するため,花序を除いた茎の最上部までの高さを測定した.ホウチャクソウについては,茎が湾曲して斜上するため,茎を伸ばした長さを測定して「高さ」とした.その他の種については,花序までを含めた高さを測定した.バイケイソウは10月には地上部が枯死していたため,春季のみ測定を行った.
なお,測定した植物のうち,フタリシズカ,バイケイソウ,シロヨメナの3種は,富士山周辺では明らかにシカが採食を避けている不嗜好植物である.
2013年6月
2013年10月
2014年10月
写真1.防鹿柵内(左)と柵外(右)の調査実施時の林床植生
2014年6月は悪天候のため撮影できなかった.
2015年5月
2015年10月
2016年6月
2016年10月
写真1.防鹿柵内(左)と柵外(右)の調査実施時の林床植生(つづき)
2.調査結果
2-1.防鹿柵内外での種組成と開花植物の違い
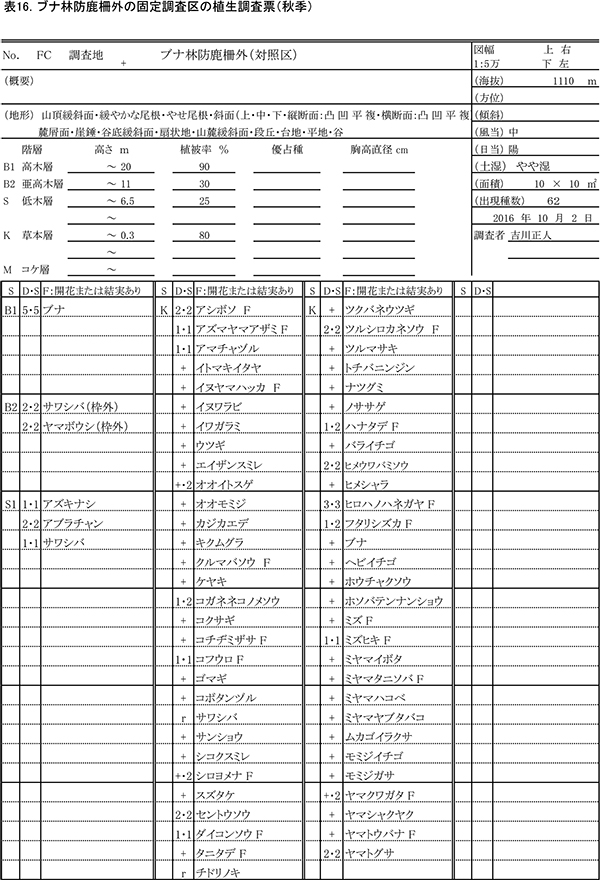
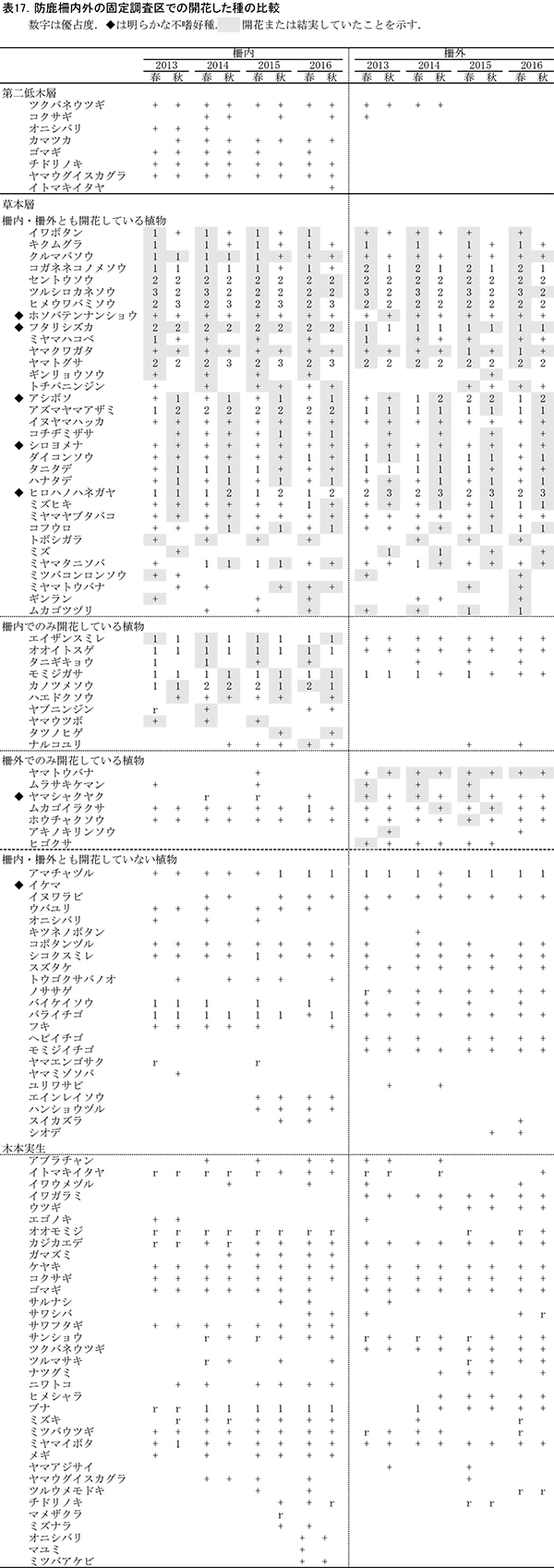
表13,14に柵内の調査区における春季と秋季の植生調査票を,表15,16に柵外の調査区の植生調査票を示す.また,表17には,2013年からの計8回の調査での出現種とその優占度を,開花の有無とともに示す.
柵内外の調査区とも,階層ごとの高さや植被率には,前回までと大きな変化はみられなかった.柵内の調査区では,前回と比べて新たに,オニシバリ,マユミ,ミツバアケビの3種の実生が出現した一方,ヤマウツボ,ヤマトウバナ,ムラサキケマンの3種は確認されなかった.柵外の調査区では,スイカズラ,ツルウメモドキが新たに出現し,ミツバコンロンソウ,アキノキリンソウなどいくつかの種が再確認されたほか,トチバニンジン,ムラサキケマン,ヒゴクサなどが消失した.しかし,全体としてみれば種組成の変化はごくわずかなものであり,優占度が大きく変化した種もなかった.
草本層の構成種の開花状況についてみると,2016年には,春秋通じて32種の植物が柵内でも柵外でも開花(または結実)していた(表17).このうち多くは,草丈が5cm未満でも開花することができる小型の植物か,茎が地表を匍匐する植物であり,草丈が少なくとも10cm以上にならないと開花しない高茎草本は,ホソバテンナンショウ,フタリシズカ,トチバニンジン,アズマヤマアザミ,イヌヤマハッカ,シロヨメナ,ダイコンソウ,ヒロハノハネガヤ,ミズヒキ,ミヤマヤブタバコの8種のみであった.また,これらのうち,ホソバテンナンショウ,フタリシズカ,シロヨメナ,ヒロハノハネガヤの4種は,まなびの森付近ではシカの採食痕跡がほとんどみられず,不嗜好植物であると考えられる.
柵内でのみ開花が観察された植物は8種あり,このうちエイザンスミレ,オオイトスゲ,タニギキョウ,モミジガサ,ナルコユリの5種は,柵外の調査区にも出現はするが,開花はしていなかったものである.ナルコユリは今回はじめて開花が確認された.一方,柵外でのみ開花が確認された植物は,昨年の調査では5種みられたが,今回はヤマトウバナ1種のみであった.
以上のように,柵内外で種組成レベルでの大きな違いは生じていないが,柵外では開花できる植物が減少する傾向が認められた.このことは,以下の種ごとの調査でも明らかであった.
2-2.防鹿柵内外での草本種のサイズと開花率の比較
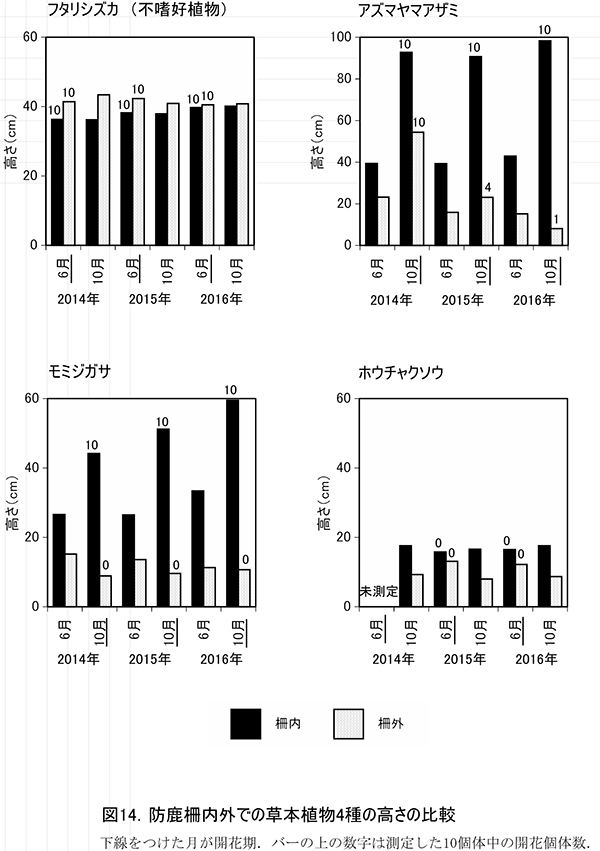
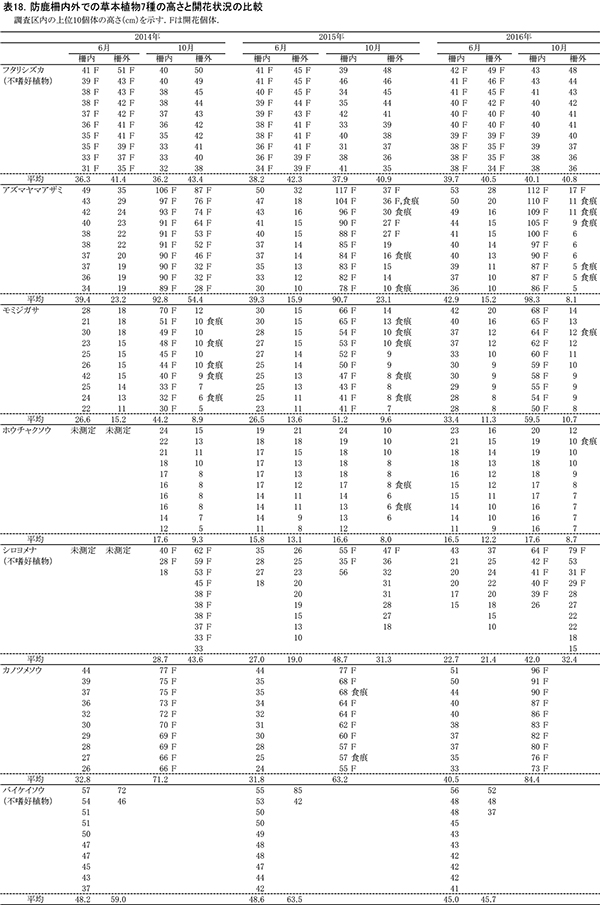
2014年から高さを測定した7種の草本種について,その測定値と開花の有無を表18に示す.このうち,毎年10個体が測定でき,柵内外での比較が可能であったフタリシズカ,アズマヤマアザミ,モミジガサ,ホウチャクソウの4種について,高さの平均値と開花個体数の変化を図14に示した.
フタリシズカは,春に茎頂に4枚の葉を展開し,その上に数本の花序をつける.花をつけた後はそれ以上伸長しないため,高さは6月,10月とも柵内では平均で約36~40cm,柵外では約41~43cmであり,3年間でほとんど変わっていなかった.また,6月には計測したすべての個体が花序をつけていた.このことから,柵外でもシカの採食をほとんど受けていないことがわかる.
アズマヤマアザミは,春から秋にかけて茎を伸ばしながら,順次葉を展開し,葉腋に花をつけていく草本である.柵内では,いずれの年も6月に40cm程度,10月に91~98cmに成長していた.しかし,柵外では6月,10月とも柵内よりも高さが低く,特に10月は柵内に比べて大幅にサイズが小さくなっていた.2016年には,10月の高さが6月よりも低く,わずか8cmまで低下しており,夏の間にシカの採食を受けたことがわかる.開花個体数も,柵内では2016年まで毎年10個体すべてが開花していたが,柵外では2015年に4個体,2016年には1個体と急激に減少した.
モミジガサは,6月頃までにほとんどの葉を展開し,秋に茎の頂部に花をつける草本である.柵内での10月の高さは年ごとに増加し,2014年の44.2cmから,2016年には59.5cmと約15cmも大きくなった.開花数も2014年以降,10個体を維持していた.一方,柵外では高さは増加せず,6月よりも10月のほうが低くなる傾向もそのままであった.開花もまったくみられなかった.
ホウチャクソウは,春季にすべての葉を展開し,6月頃までに開花する植物である.モミジガサと同様,柵外では6月よりも10月の高さが低くなっており,シカに採食されていることがわかる.柵内でもまだ高さの回復はみられず,開花個体も観察されなかった.
以上のように,柵の設置に対する草本種の反応は,種によってさまざまであった.これらの反応の違いからは,それぞれの種が柵の設置までに受けてきた採食圧の影響と,柵によって保護されたときの回復力の違いを読み取ることができる.柵内でのサイズや開花率が変化せず,柵外でのサイズの低下が大きかったアズマヤマアザミは,柵を設置した2014年の時点では,採食圧の影響がほとんど現れていなかったと考えられる.この時点では,アズマヤマアザミはあまり採食されていなかったか,食べられていてもサイズや開花率に影響を及ぼすほどではなかったが,その後も採食圧がかかり続けた柵外では,この3年間で急速に成長の抑制と開花率の低下が生じたといえる.つまり,今回の柵設置のタイミングは,アズマヤマアザミにとっては,採食の影響が出始める直前であったと考えられる.
対照的に,モミジガサは柵内のサイズや開花率が大幅に回復している.この種は,柵を設置した2014年までに,繰り返し採食されたことによる地下の貯蔵器官の消耗が激しく,地上部のサイズが小さくなっていたが,柵で保護されたことによって回復したとみなされる.柵外では,いまでも採食圧がかかり続けているために,高さや開花率は元のままである.モミジガサは,柵設置の時点ですでに大きな採食影響を受けていたものの,採食圧から解放されれば,すみやかに回復できる程度のダメージであったと考えられる.
また,柵設置後もサイズや開花に変化がみられないホウチャクソウは,モミジガサに比べて採食の影響が強く,回復に時間がかかるのかもしれない.これらの結果から,①柵を設置した以外の場所では採食圧がかかり続けており,これまで影響を受けていなかった植物にも影響が出ていること,②柵の設置によって一部の植物は本来の生育状態を回復したが,3年程度では回復できないダメージをすでに被っている種もあること,が明らかになった.現在まで,柵の有無によって種組成レベルの違いは生じていないが,将来的にはシカが好む植物の個体の小型化や繁殖率の低下,不嗜好植物が相対的に有利になることなどによって,種組成にも違いが生じてくることが予想される.


「まなびの森」の20年の活動をまとめたPDFです
![]()