








































モニタリング活動
植生調査 2017年度報告書
1.はじめに
富士山南麓では1996年(平成8年)9月の台風17号により大規模な風倒被害が発生し,壮齢ヒノキ林を中心に約90,000㎥が被害を受けた.富士山「まなびの森」事業計画では,この風倒被害跡地に広葉樹を中心とした在来樹種を植栽することによって,早期に自然林を復元することを目指している.本調査は,「まなびの森」区域内での台風による風倒跡地の森林の回復および変化についてモニタリングをおこない,自然林の早期再生のための管理・施業方法に関する情報を得ようとするものである.
2017年度は,以下の2点についてモニタリング調査を実施した.
1)風倒被害跡地の群状植栽地および未植栽地との比較対象として2000年に設置した,ブナ自然林,ヒノキ若齢林,ヒノキ壮齢林,ウラジロモミ林,人工林風倒ギャップの5つの固定調査区について,樹木の生育状況と種組成に関する追跡調査をおこなった.この結果を過去3回の調査結果と比較することによって,17年間の植生変化を明らかにした.
2)2013年にブナ自然林内に設置した防鹿柵内外の固定調査区について,林床の種組成に関する追跡調査を行った.この結果を過去4年間の調査結果と比較することによって,「まなびの森」周辺に多数生息するニホンジカが,森林植生に与えている影響を明らかにした.
2.調査地と方法
2-1.森林の調査区
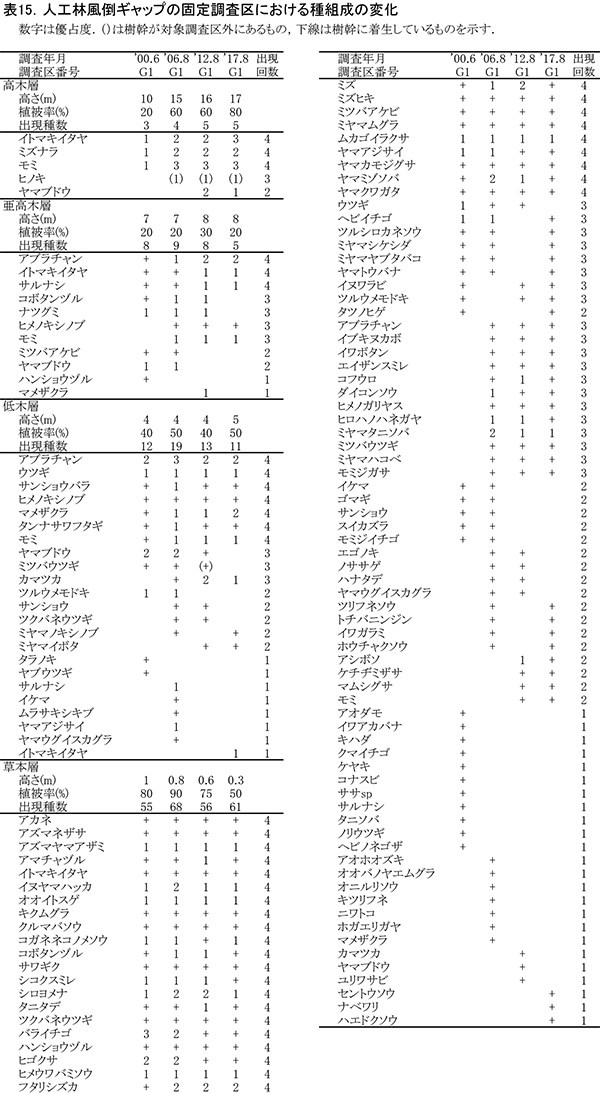
植栽地および未植栽地との比較のために森林内に設けた,5ヶ所の固定調査区(図1)では, 2001年,2006年,2012年の3回にわたって,5-6年おきに毎木調査と種組成の調査が実施されている(初回の種組成の調査のみ2000年).2017年は4回目の調査となる.各調査区の概要は以下のとおり.
①ブナ自然林(調査区番号F1~9,900㎡)
富士山の南麓斜面を代表するブナ林の典型的な部分に設置したものである.直径60cmを超えるブナとケヤキの大径木が林冠を占めており,その下層にミズキ,サワシバ,チドリノキ,エンコウカエデ,ヤマボウシ,アブラチャンなどが生育する林分である.林床には周辺地域で繁茂することが多いスズダケが生育せず,かわりに多数の草本種が生育する.人工林に比べて林冠を構成する高木がまばらであるため,10m×10mの調査区を9つ組み合わせた30m×30mの調査区とした.
②ヒノキ若齢林(調査区番号Cy1,100㎡)
1982年に植林された林分で,調査開始時の林齢は18年,現在35年生のヒノキ林である.調査区内には,現在生育しているヒノキより直径の大きな切り株が残されていることから,2世代目の人工林であることがわかる.調査区内では2010年に間伐がおこなわれた.また,2011年夏の大雨により,調査区内に土砂が流入した.
③ヒノキ壮齢林(調査区番号Co1~2,200㎡)
1958~1959年の台風による風倒被害後,1960年に植栽された林分で,調査開始時の林齢は40年,現在57年生のヒノキ林である.調査開始以前に一度間伐がおこなわれており,樹木密度は若齢林よりも小さい.前回の調査後,2014年頃にも間伐が実施された.10m×10mの調査区を斜面方向に2つ組み合わせて10m×20mの調査区とした.
④ウラジロモミ林(調査区番号A1,100㎡)
1945年に植林された林分で,調査開始時の林例は55年,現在72年生のウラジロモミ林である.2001年の時点で,生存していたウラジロモミの17本のうち10本の根元部分にシカによる剥皮に被害がみられ,調査開始前からシカの食害の影響を受けてきた.2011年夏の大雨により,調査区内に土砂が流入した.
⑤人工林風倒ギャップ(調査区番号G-1,100㎡)
1996年の台風時に風倒によって生じたギャップの回復過程を追跡するために設置したものである.この付近は1964年にウラジロモミ,ヒノキ,シラベが混植された人工林であるが,調査区を設置した場所では主にモミが生育していた.風倒被害が発生したときの林齢は32年で,直径20~25cm程度のモミ3本が折れたために生じたギャップである.
これらの調査区において,2017年8月6日~9日にかけて,次の調査を行った.
1)毎木調査
調査区内に生育している胸高直径(以下,DBH)5cm以上の樹木について,樹高およびDBH を測定した.樹高は検測棹をもちいて10cm単位(10m以上は目測),DBHは直径巻尺をもちいて㎜単位で計測した.また,個体識別のためにナンバーテープをつけ,位置図と樹冠投影図を作成した.
2)植生調査
10m×10mの区画を単位として,植物社会学的な方法による植生調査をおこなった.これは,出現するすべての植物をリストアップして,それぞれの種の優占度および群度を判定するものである.優占度・群度の階級はBraun-Blanquet(1964)の判定基準にしたがい,優占度はr,+,1~5の7段階,群度は1~5の5段階で記録した.
2-2.防鹿柵内外の固定調査区
2013年3月に,シカの採食から林床植生を保護する目的で,ブナ自然林の固定調査区から約50m東側の緩斜面に,およそ13m×13m程度の防鹿柵を設置した(図1).ネットはステン線入りナイロン製,目開き径5cmで,高さ2mで支柱間隔は約3mである.この内部に10m×10mの固定調査区を設置した.また,防鹿柵の効果を検証するための比較対象として,柵外の近接する場所に同じサイズの対照区を設置した.調査区番号はFN(防鹿柵内)およびFC(防鹿柵外)である.これらの調査区では,2013年以降,年2回(春季:5月末~6月始め,秋季:9月末~10月始め)植生調査を実施している.2017年は5年目の調査となる.
これらの調査区において,2017年6月1日および9月29日に,以下の調査を行った.
1)植生調査
森林の調査区と同様の方法で,植物社会学的な植生調査を行った.草本種については,開花・結実の有無も記録した.
2)主な草本種の草丈の測定
主要な草本種のサイズや開花率に対するシカの採食の影響を知るため,調査区内の代表的な高茎草本7種(フタリシズカ,アズマヤマアザミ,モミジガサ,ホウチャクソウ,カノツメソウ,バイケイソウ,シロヨメナ)について,調査区内の上位10個体の高さと開花の有無,食痕の有無を記録した.これらのうち,フタリシズカについては,茎の頂部につく花序の有無が高さに影響するため,花序を除いた茎の最上部までの高さを測定した.ホウチャクソウについては,茎が湾曲して斜上するため,茎を伸ばした長さを測定して「高さ」とした.その他の種については,花序までを含めた高さを測定した.
なお,測定した植物のうち,フタリシズカ,バイケイソウ,シロヨメナの3種は,明らかにシカが採食を避けている不嗜好植物である.
3.調査結果
3-1.森林の調査区における過去17年間の変化
3-1-1.ブナ自然林
(1)樹木の生育状況
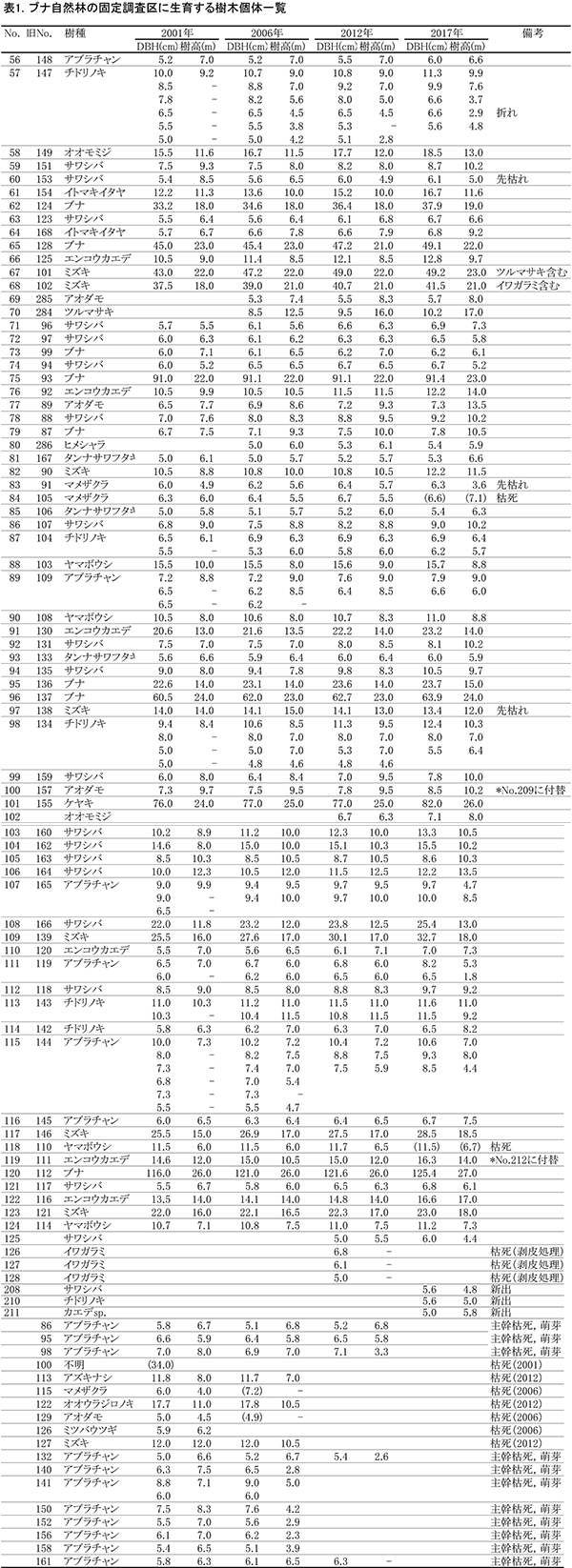
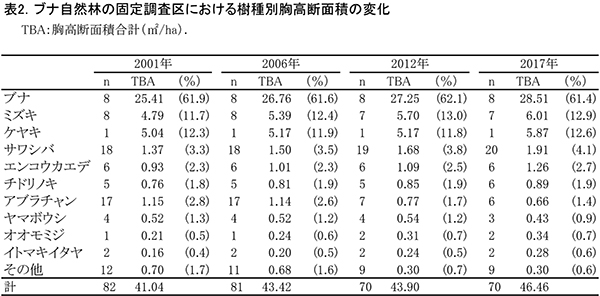
表1に調査区内に生育する樹木の胸高直径と樹高の一覧を,表2に樹種別の胸高断面積合計値を示す.
2006年から6年間の間に,DBH6.7cmのヤマボウシ(No.118)と,DBH7.1cmのマメザクラ(No.84)が枯死していた.また,アブラチャン5個体がDBH5cm以上の幹が枯死して,測定対象外となった(表1).アブラチャンの枯死は2006年の調査時から続いており,個体数は調査を開始した2001年の17個体から6個体まで減少した(表2).ただし,これらの株からは細い萌芽幹が再生しており,個体としては枯死してはいないものが多い.枯死したアブラチャンの幹にはシカによる樹皮剥ぎの痕は認められず,食害による枯死ではないと考えられる.おそらく,直径7-8cmを超えると肥大成長の限界を迎えて,新しい萌芽と主幹を交代させる特性をもっているのであろう.ブナの林冠木にからみついていたイワガラミ3個体は,人為的に剥皮処理がなされて枯死していた.一方,2017年は新たにサワシバ,チドリノキ,カエデ属の1種の新出個体がみられた.
樹種別の本数と胸高断面積合計(TBA)の割合は,アブラチャンの減少を除いて,2001年以降大きく変化していない(表2).ブナが全樹種の約62%を占め,ミズキとケヤキが約13%ずつで続いていた.亜高木層以下では,個体数・TBAともサワシバが最も多く,20個体で約4%を占めていた.樹種の数としては,オオウラジロノキ,アズキナシ,ミツバウツギが期間中に枯死したため,3種減少していた.
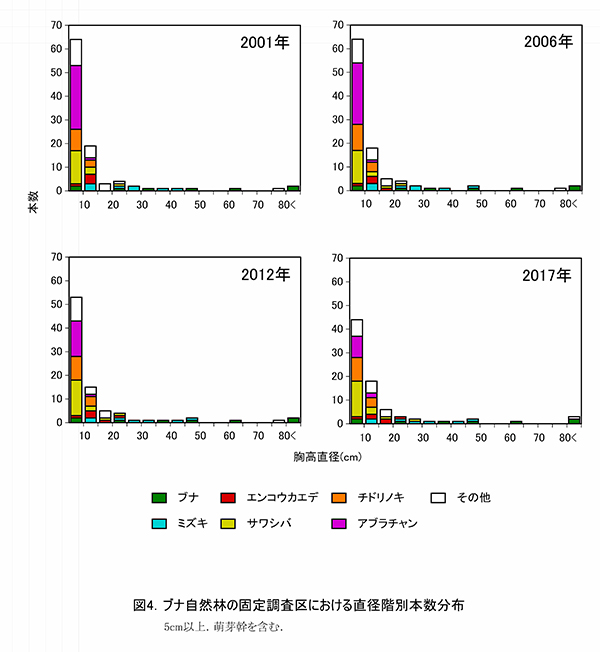
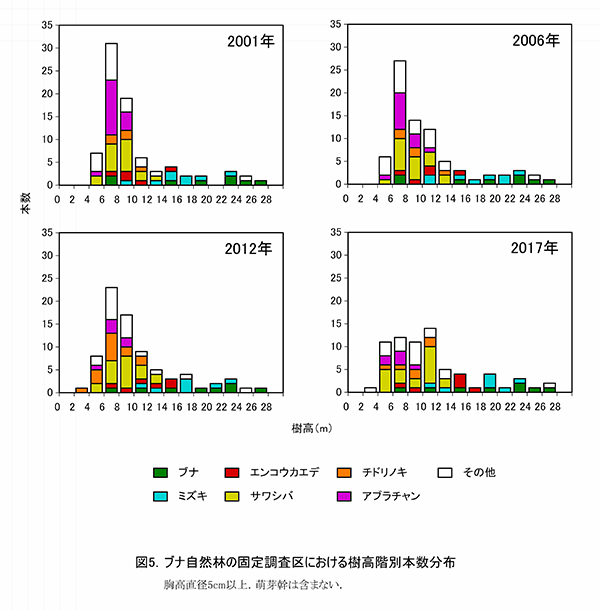
図12,13に直径階別および樹高階別本数分布の経年変化を示す.直径階別本数分布をみると,5‐10cmに本数のピークがあり,50cmまでは連続的に本数が減少してL字型の分布をしている.林冠を占めるブナ3個体とケヤキ1個体は,突出して直径が大きな個体であることがわかる(図12).2001年,2006年と比較すると,分布型に大きな変化はないが,アブラチャンの枯死により,5‐10cmの本数が減少している.樹高階別本数分布でも同様に,6‐8mにピークがあることは変わりないが,その本数はやや減少していた.
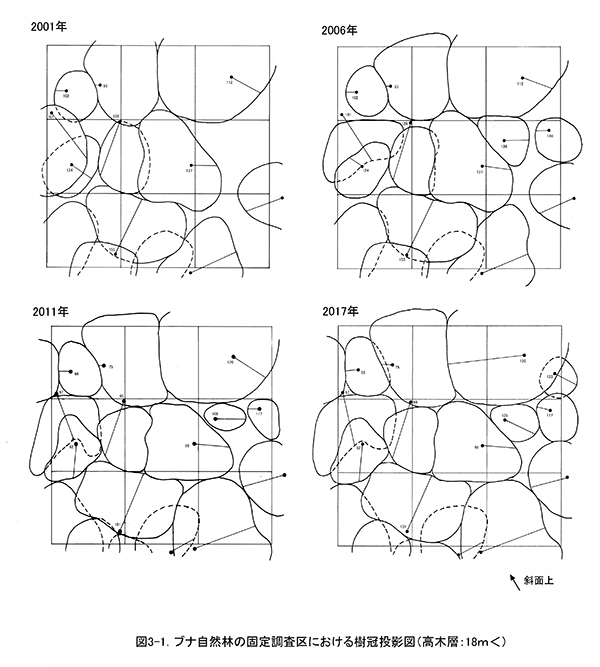
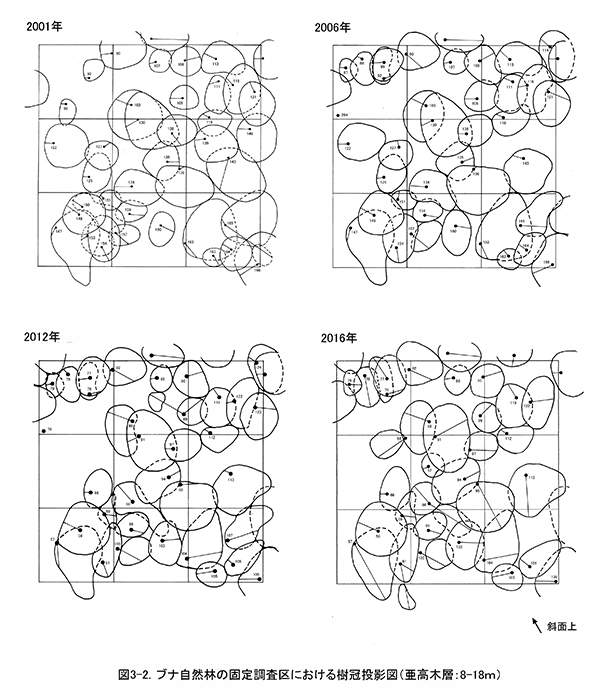
図2は調査区内の樹木の位置,図3に階層別の樹冠投影図である.樹冠投影図は複雑になるので,高木層(おおむね18m以上),亜高木層(8~18m),低木層(8m未満)の3階層に分けて示した.
調査区内の林冠は,ブナ,ケヤキ,ミズキと,調査区の下方から樹冠を広げているオニイタヤによって占められている.2001年当初は高木層にやや隙間があったが,2006年以降はミズキ(No.109,117,123)が徐々に樹高を増して高木層に加入し,現在はほぼ林冠がふさがった状態になっている(図3-1).亜高木層もよく発達しており,前述のように数個体の枯死はみられたが,全体としては樹冠面積が増加している(図3-2).これに対して,低木層はややまばらで,分布に偏りがある.2006年以降のアブラチャンの枯死により,斜面下方を中心に低木層を欠く範囲が広がってきた(図3-3).
図4に直径階別,図5には樹高階別の本数分布を示す.直径階別本数分布は逆J字に近い形を示し,林冠を構成しているのはDBH30cm以上の樹木である.このうちブナ2個体とケヤキ1個体がDBH80cmを超えている.DBH15cm未満の小径木には,サワシバ,チドリノキ,アブラチャンが多い(図4).直径階別本数分布が逆J字型を示すのに対し,樹高階別本数分布の図は4m未満の個体が少ない一山型の形状を示し,低木層の貧弱さが表れている(図5).亜高木層の構成種の中では,エンコウカエデとサワシバに樹高が大きいものがみられる.樹高8m未満のブナの幼樹は2個体しかみられず,現在の林冠を構成している大径木が枯死した後は,ブナ林としては維持されず,カエデ類を中心とした樹高の低い森林群落に移行していく可能性がある.
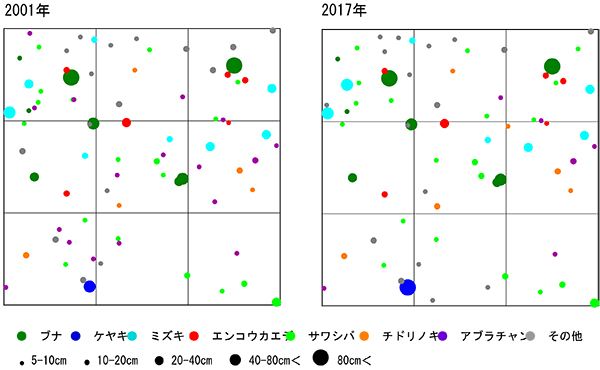
図2.ブナ自然林位置図
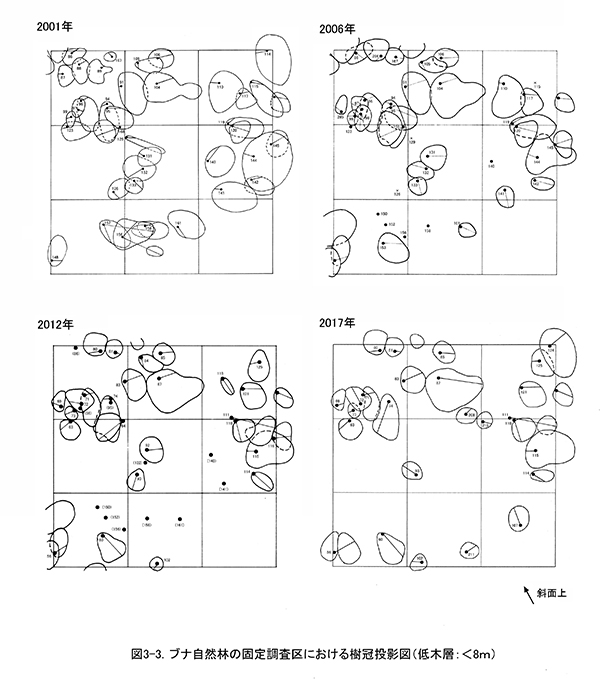
(2)種組成
ブナ自然林の固定調査区における階層ごとの出現種とその優占度の変化を表3に示す(個別の植生調査票は別冊,付表1~9).2000年以降,高木層と亜高木層では,ミズキの一部の個体が亜高木層から高木層に加入したこと,亜高木層のオオウラジロノキやアズキナシが枯死したことのほか,大きな変化はみられなかった.しかし,低木層から草本層にかけては構成種に明らかな変化がみられた.
低木層では,2006年から2012年の間に,ミツバウツギ,コクサギ,マメザクラ,カマツカといった樹種が消失していた.調査区あたりの平均出現種数も,2000年の6.7種,2006年の6.2種から,2012年は3.4種に大きく減少した.2012年までに消失した樹種は2017年になっても回復しておらず,平均出現種数も3.6種と少ないままである.前述のアブラチャンの枯死もあり,2006年以降は低木層の衰退が進んでいる.
草本層では,調査区あたりの平均出現種数は2000年の49.8種から,2006年は47.4種,2012年は43.6種,2017年は48.0種と,顕著な減少はみられない.しかし,植被率は2000年に50~80%の範囲にあったのが,2017年には20~40%に低下していた.落ち葉が分解して少なくなる夏季は,草本の植被のない部分では土壌がむき出しになり,雨水による浸食をうけつつある場所もみられる.
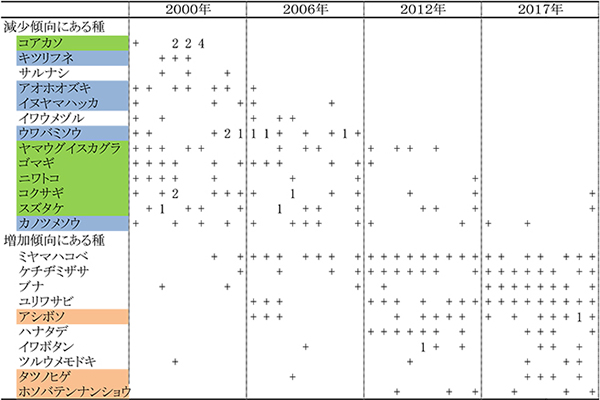
出現頻度に明らかな変化があった種を,表3の「草本層」の上部に配置してある.2000年に3つ調査区で優占度2以上で生育していた低木のコアカソは,2006年にはまったくみられなくなり,その後も回復していない.このほか,キツリフネ,サルナシが2006年までに,アオホオズキ,イヌヤマハッカ,イワウメヅルが2012年までに消失した.さらに,ウワバミソウ,ヤマウグイスカグラ,ゴマギが2017年までに消失した.そのほか,出現頻度は変わらないが優占度が低下する傾向がある種として,ムカゴイラクサ,アズマヤマアザミ,オオイトスゲなどが挙げられる.
一方,2000年には記録されなかったが,2006年以降出現した種には,ユリワサビ,アシボソ,ハナタデ,タツノヒゲなどがある.また,ミヤマハコベやケチヂミザサは出現頻度が大幅に増加した.さらに,フタリシズカ,セントウソウ,コガネネコノメソウ,ツルシロカネソウ,シロヨメナなどは,優占度の増加傾向が認められた.草本層で出現頻度や優占度が増加している種は,シカがあまり採食しない不嗜好性植物(アシボソ,タツノヒゲ,フタリシズカ,シロヨメナなど)であるか,植物体が小さいために採食されにくい植物(ミヤマハコベ,セントウソウ,コガネネコノメソウ,ツルシロカネソウなど)が多い.草丈の高い草本植物がシカの採食によって減少する一方で,シカが食べない,あるいは食べられにくい草本植物が増加する傾向が顕著であった.
以上のように,2000年以降,森林の階層構造と林床の種組成に大きな変化が生じたことが明らかになった.すなわち,2012年までに低木層が著しく衰退して「ディアライン」が形成されるとともに,高茎草本の減少が顕著になり,不嗜好植物や小型の草本種によるまばらで丈の低い林床に変化した.「まなびの森」を含む富士山南麓域では,1990年代からシカの生息密度が急増し,調査を開始した時点で,すでにシカの採食圧の影響を受けていたが,2000年代に入って採食圧の蓄積が植物の耐性の限度を超え,階層構造や種組成の変化として顕在化してきたといえる.ただし,2006年以降は,低木や高茎草本が減少してシカの餌場としての利用価値が低下したためか,地表に落ちているシカの糞の量が減少したように観察される.
表3.ブナ林組成表
3-1-2.ヒノキ林(若齢)
(1)樹木の生育状況
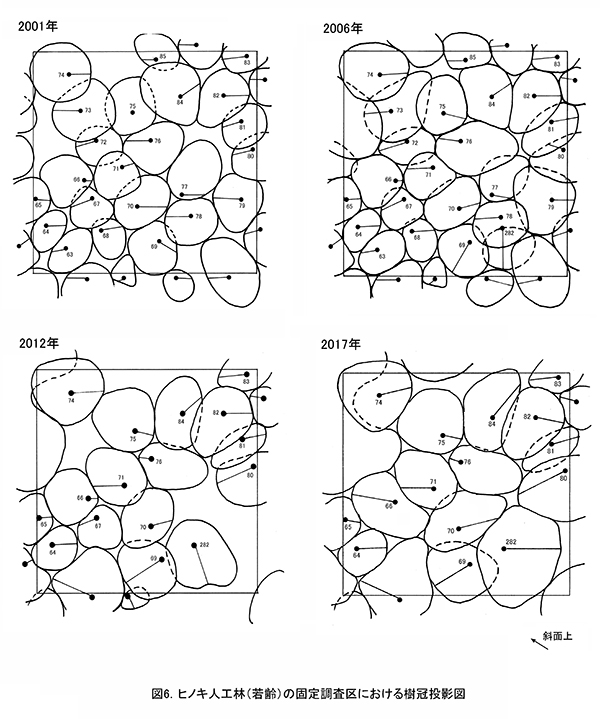
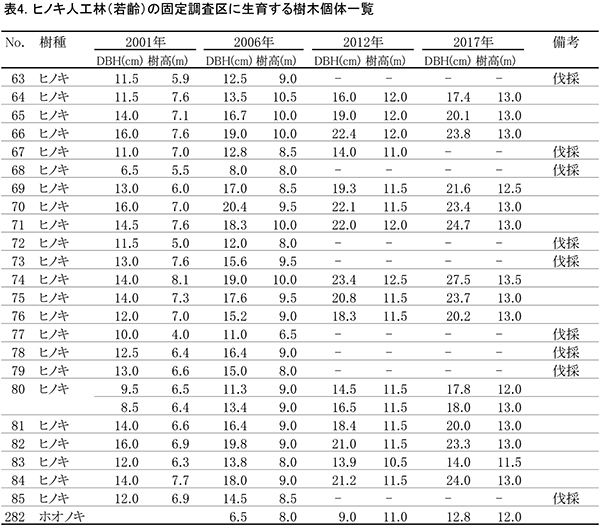
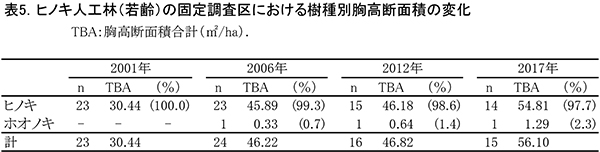
表4に調査区内に生育する樹木の胸高直径と樹高の一覧を,表5に樹種別の胸高断面積合計値を示す.また,図6には調査区内の樹冠投影図,図7,8にはそれぞれ胸高直径階別,樹高階別の本数分布を示す.
2001年の調査時には23本のヒノキが生育していたが,2010年に間伐が行われて8本が伐採された.その後,生育不良の1本も伐採されたため,2017年に生育しているヒノキは14本である.樹冠投影図の変化をみると,間伐により生じたギャップは,2017年までにほとんど埋められていることがわかる(図6).また,No.282のホオノキと左側枠外から伸びているヤマグワの樹冠が大きく拡大した.ヒノキの胸高断面積合計は2001年の30.4㎡/haから2017年には54.8㎡/haに大きく増加した(表5).
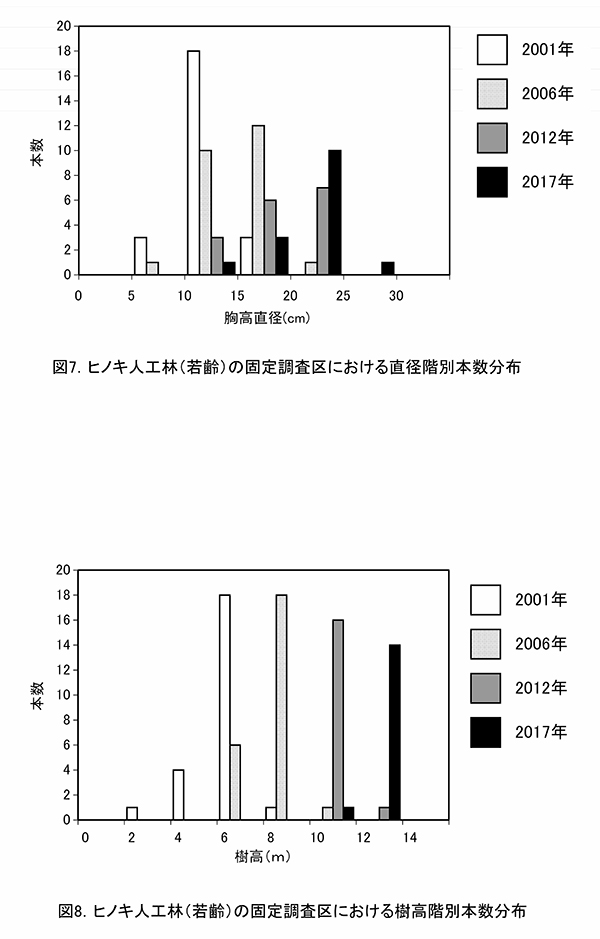
ヒノキの胸高直径階別本数分布の経年変化(図7)をみると,2001年から最も本数の多い階級が5cmずつ移動し,現在では20~25cmの階級で最も本数が多くなっていた.樹高階別本数分布(図8)でも,2001年以降,最も本数が多い階級が2mずつ移動し,現在は12~14mの階級で最も多くなっていた.適切な間伐によって良好な成長が維持されているといえる.
(2)種組成
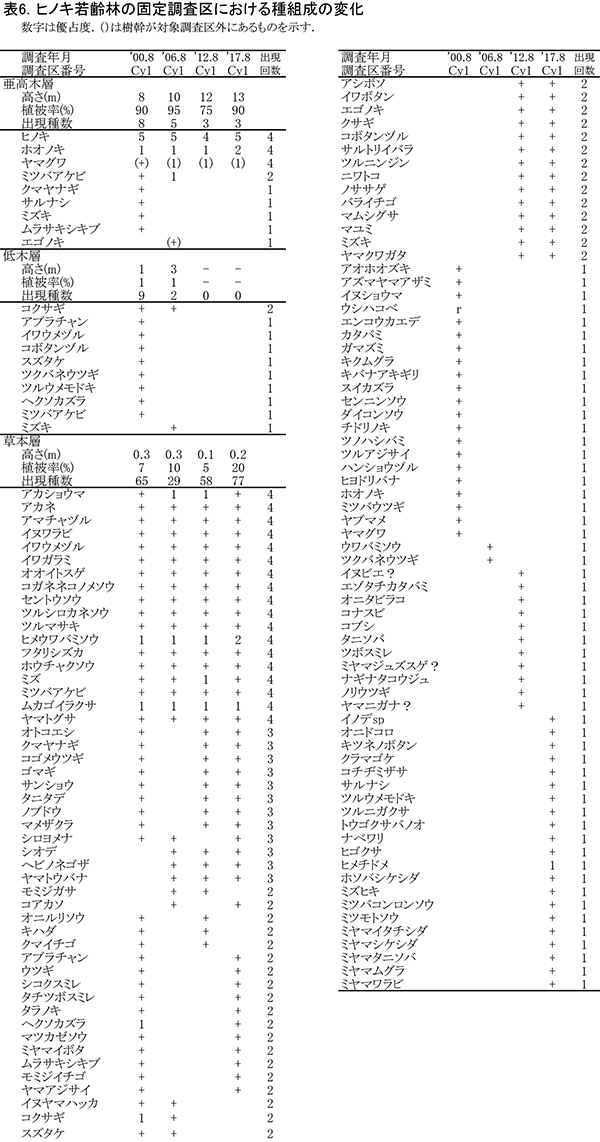
階層ごとの出現種とその優占度の変化を表6に示す(植生調査票は別冊,付表10).調査開始時の2001年には,林冠が閉鎖していなかったため,亜高木層から低木層にはムラサキシキブ,コクサギ,アブラチャンなどの低木性樹種がみられるとともに,亜高木層にはミツバアケビ,クマヤナギといったつる性木本がヒノキにからみついていた.林冠が閉鎖した2006年以降は,これらはほぼみられなくなり,間伐にともなう管理の影響もあって,2017年には亜高木層と草本層のみからなる単純な階層構造になっている.
草本層の出現種は調査年によってばらつきが大きく,4回の調査を通じて出現したのは18種のみであった.このうち比較的優占度が大きいのは,アカショウマ,ヒメウワバミソウ,ムカゴイラクサであった.2001年には草本層の出現種数は65種であったが,林冠が閉鎖した2006年には29種にまで減少した.しかし,間伐後の2012年には再び58種に増加している.このときは間伐に加えて,2011年に大雨による土砂の流入が生じるなど,地表の攪乱が大きかったためか,オニタビラコ,コナスビといった森林生でない雑草的な草本種の出現もみられた.2017年にはさらに出現種が増加して77種にも達した.構成種にもトウゴクサバノオ,ナベワリなど,周辺のブナ自然林を特徴づけるものが出現している.
3-1-3.ヒノキ林(壮齢)
(1)樹木の変化
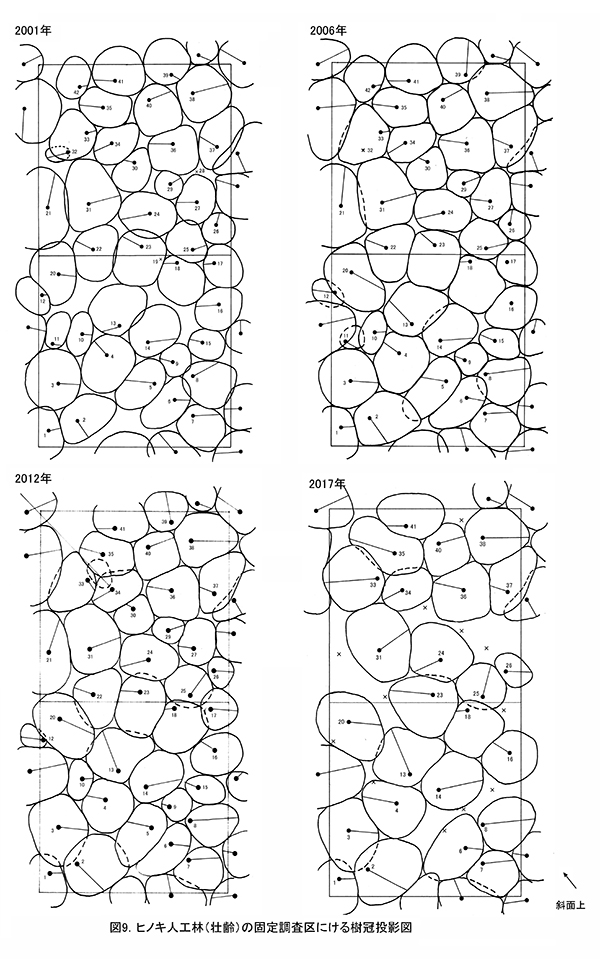
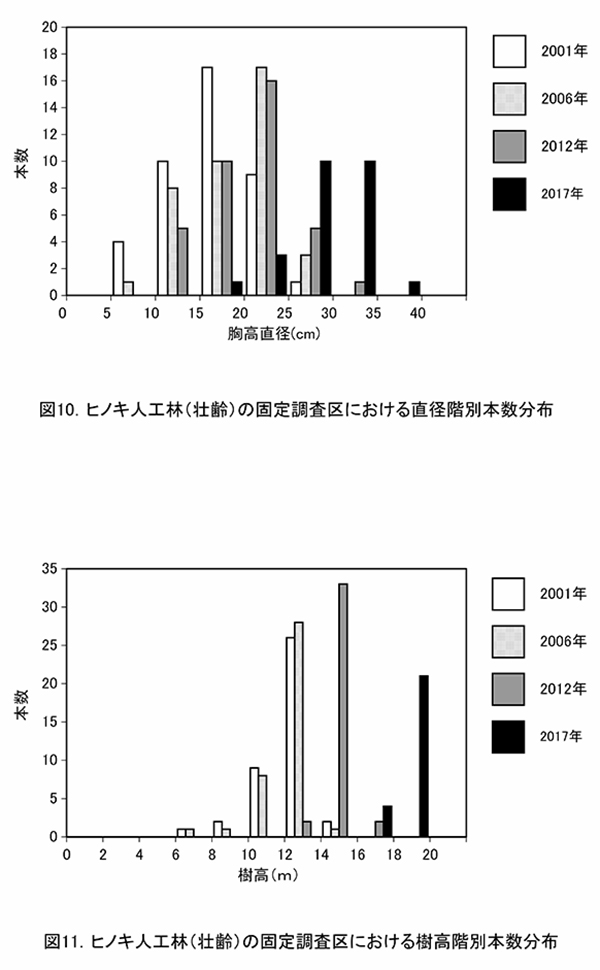
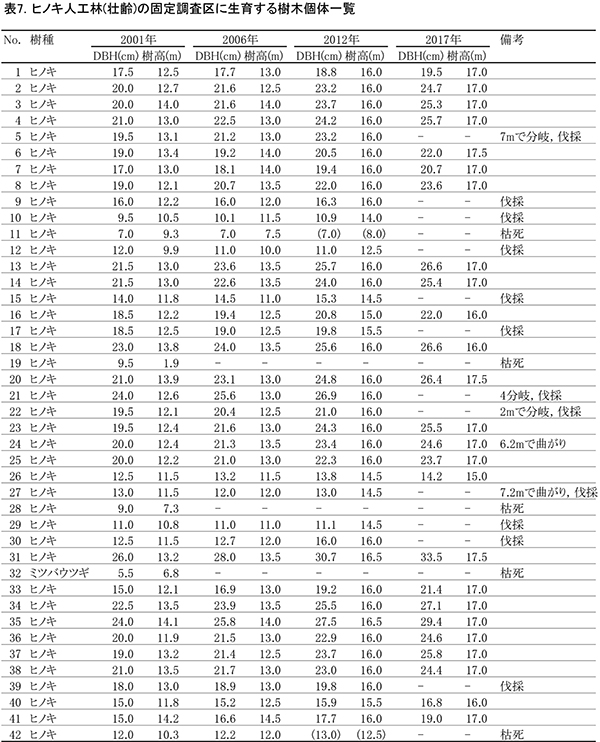
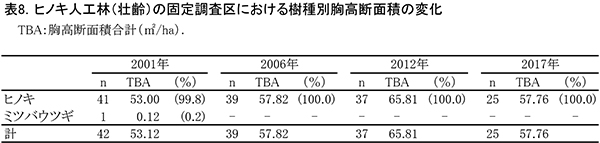
表6に調査区内に生育する樹木の胸高直径と樹高の一覧を,表7に樹種別の胸高断面積合計値を示す.また,図9には調査区内の樹冠投影図,図10,11にはそれぞれ胸高直径階別,樹高階別の本数分布を示す.
2001年の調査開始時には41本のヒノキが生育していたが,2012年までに4本が枯死して37本となった.その後間伐が行われ,2017年に生育していたヒノキは25本である(表7).2006年から2012年にかけて,隣接個体どうしの樹冠が密に接した状態であったが,2017年には間伐で樹冠面積の小さな個体が除かれ,空隙ができた状態になっている.胸高断面積合計は,2012年に65.8㎡/haであったが,間伐後の2017年には2006年とほぼ同じ57.8㎡/haとなっている(表8).
ヒノキの胸高直径階別本数分布の経年変化(図10)をみると, 2012年までDBH20~25cmの階級で最も本数が多かったが,2017年になると25~30cm,30~35cm以上の階級に属する本数が増加した.また,樹高階別本数分布(図11)では,2012年には14~16mに属する個体が最も多かったが,2017年には2階級上の18~20mにほとんどの個体が移行していた.これらから,2012年以降に行われた間伐の効果によって,樹木の生長速度が増大したことが読み取れる.
(2)種組成の変化
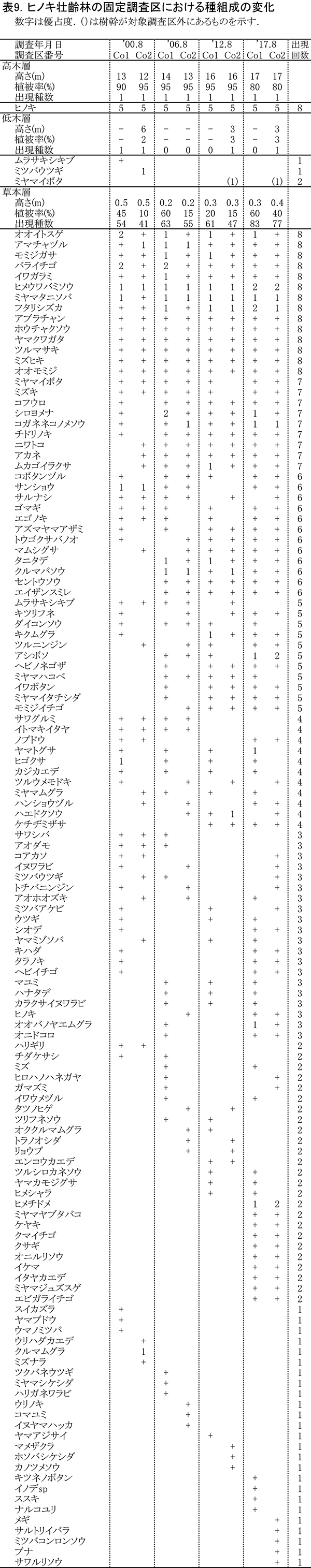
階層ごとの出現種とその優占度の変化を表9に示す(植生調査票は別冊,付表11~12).ヒノキ若齢林と比べて,当初から林冠が閉鎖していたため亜高木層と低木層は発達せず,高木層と草本層からなる単純な構造である.
草本層の植被率は,斜面下側の風倒地に隣接する方形区Co-1のほうが,斜面上側の自然林に接するCo-2よりも高い.Co-1では,2012年に60%から20%に低下したが,間伐後の2017年には再び60%に回復した.草本層の構成種のうち比較的優占度が高いのは,オオイトスゲ,ヒメウワバミソウ,ミヤマタニソバ,フタリシズカなどである.経年的な変化をみると,サワグルミ,サワシバ,アオダモといった高木性樹種の実生が2012年以降みられなくなったこと,タニタデ,クルマバソウ,セントウソウ,エイザンスミレなど周辺のブナ自然林に多い種が2006年以降に出現し始めたことがわかる.さらに2017年には不嗜好植物であるアシボソの優占度が増加するとともに,これまで記録されなかったミヤマヤブタバコが優占度1または2で出現した.ヒノキ若齢林の調査区に比べると,ヘビノネゴザ,ミヤマイタチシダといったシダ植物が多く出現していた.これらは,間伐後その場に残置された木材の隙間が生育場所となっていることが多い.
なお,2017年には,山梨県,神奈川県など近県で絶滅危惧種とされており,「まなびの森」では初記録となるサワルリソウが確認された.
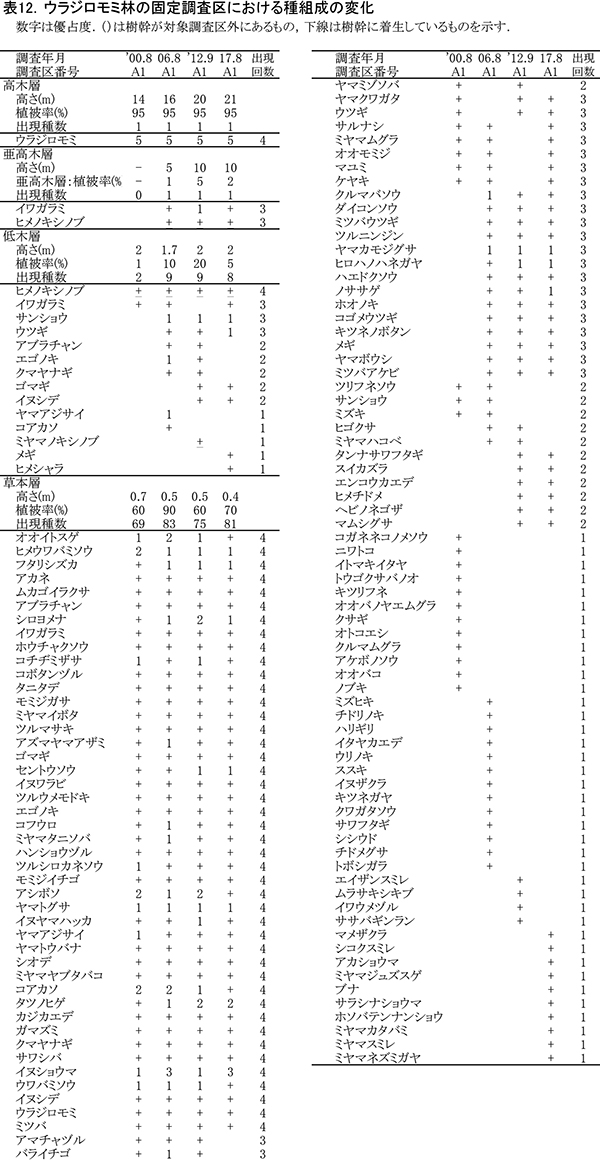
3-1-4.ウラジロモミ林
(1)樹木の変化
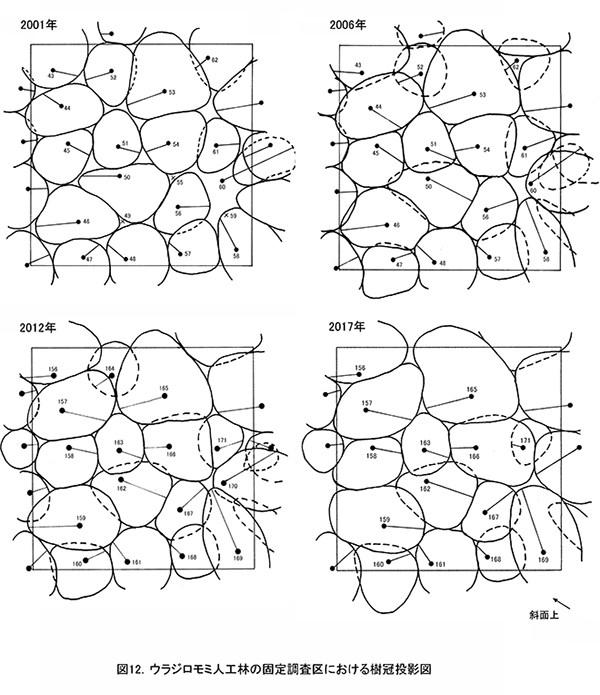
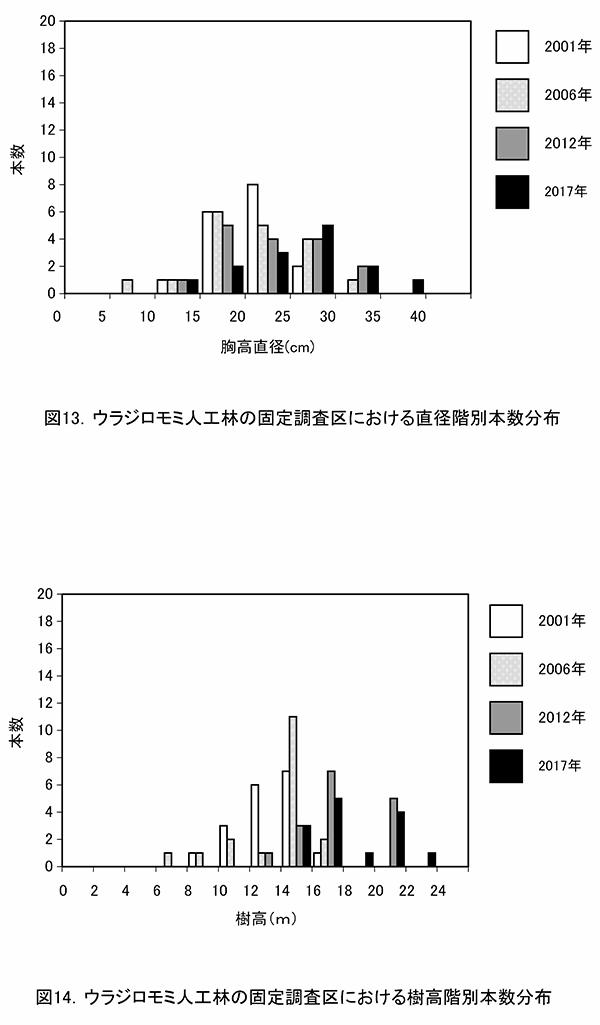
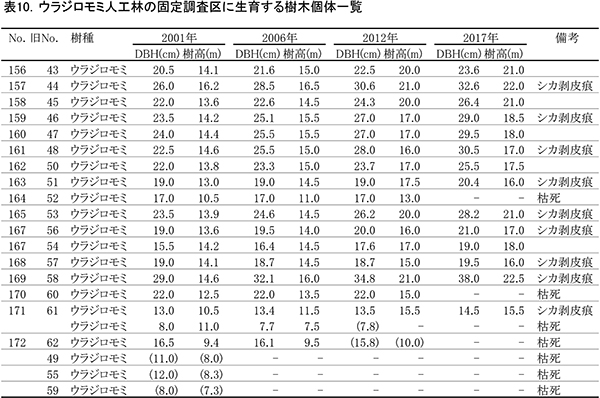
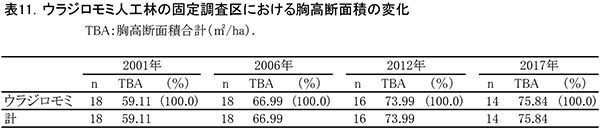
表10に調査区内に生育する樹木の胸高直径と樹高の一覧を,表11に樹種別の胸高断面積合計値を示す.また,図12には調査区内の樹冠投影図,図13,14にはそれぞれ胸高直径階別,樹高階別の本数分布を示す.
ここでは調査期間を通じて間伐は行われていないが,当初生育していた18本のウラジロモミのうち4本が枯死し,2017年までに14本になった.調査を開始した2001年までに枯死していたものも3本あり,個体間の競争による間引きが進んできたと考えられる.調査区内のほとんどのウラジロモミには,根元部分にシカの採食による剥皮痕がみられた(表10)が,全周を剥皮されたものはなく,枯死の直接の原因は剥皮よりも周囲の個体による被圧と考えられる.胸高断面積合計は2001年の59.1㎡/haから,2012年には74.0㎡/haに増加したが,2012年以降は増加量が小さく,2017年のTBAは75.8㎡/haであった(表11).
間伐が行われていないため,樹冠面積や直径・樹高階別の本数分布は,前述のヒノキ林の2調査区に比べるとばらつきが大きい.樹高階別本数分布をみると,2006年には14~16mの個体が多く樹高がそろっていたのに対し,2012年になると18~20mと22~24mの2つの階級にピークができ,この傾向が2017も続いている.樹高成長の個体差が大きくなり,上層木に被圧される個体がでてきているといえる(図14).TBAの増加が頭打ちになっていることからみても,樹木が過密になっており,間伐が必要な状態になっていることがわかる.
(2)種組成の変化
階層ごとの出現種とその優占度の変化を表12に示す(植生調査票は別冊,付表13).ここでは,2001年にはほとんど低木層が認められなかったが,2006年にから2012年にかけては,サンショウ,ウツギ,エゴノキなどがわずかながら低木層を形成していた.これは,調査区の斜面下側で浸食谷の形成が進み,倒木が生じて林内に光が当たるようになったことが原因と考えられる.しかし,2017年には再び低木層の植被率が20%から5%に低下していた.
草本層の植被率も2006年には一時的に90%まで増えたが,その後は60~70%で推移している.ここでは,主要な構成種は,他の調査区にも多いオオイトスゲ,ヒメウワバミソウ,フタリシズカなどのほか,イヌショウマが目立つ.ブナ自然林で2012年までに消失したコアカソは,ここでも優占度2から+まで減少した.また,アシボソは不嗜好植物で他の調査区では増加傾向にあるが,ここでは光環境の悪化のためか,2017年には優占度2から+に減少していた.不嗜好性植物のフタリシズカ,シロヨメナ,タツノヒゲ,ヤマカモジグサ,ヒロハノハネガヤなどは2006年以降,優占度が増加する傾向にある.
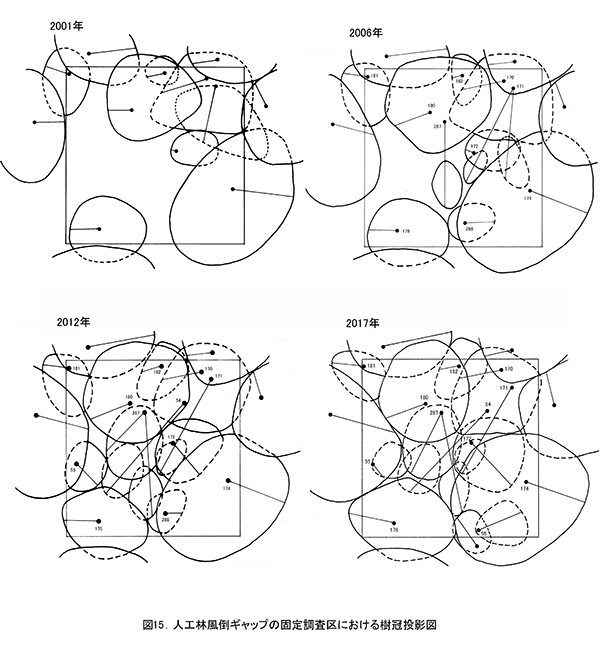
3-1-5.人工林風倒ギャップ
(1)樹木の生育状況
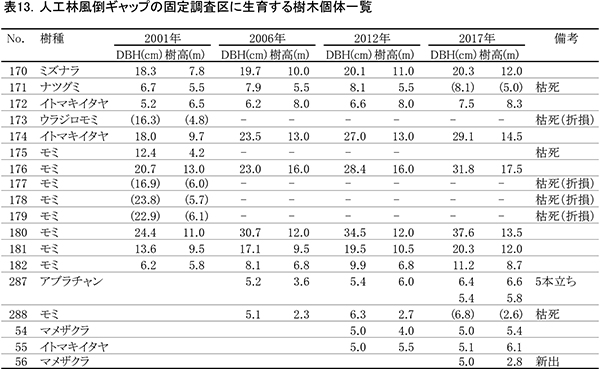
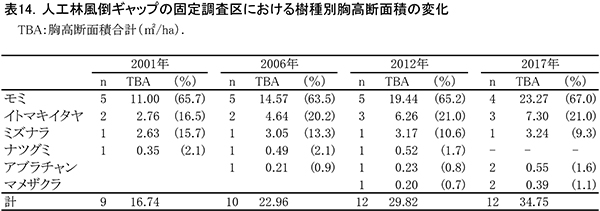
表13に調査区内に生育する樹木の胸高直径と樹高の一覧を,表14に樹種別の胸高断面積合計値を示す.また,図15には調査区内の樹冠投影図を示した.
ここは林冠を構成していたDBH20cm前後のモミが台風で折れてできた,直径5mほどのギャップである.このギャップの部分には高木の稚樹や低木性樹種は生育していなかったが,2012年までに周囲からアブラチャン,ナツグミ,マメザクラなどの低木が極端に斜上する幹を伸ばした.また,これらの低木の上には,ヤマブドウ,ツルウメモドキといったつる性木本が繁茂して,傷口をふさぐようにギャップを埋めていった.さらに風倒を免れたモミ(No.176,180),イトマキイタヤ(No.174)の樹冠が拡大することによって,その上層もふさがれつつある(図15).これらの直径は過去16年間で大きく増加し,No.176のモミとNo.174のイトマキイタヤは約11cm,No.180のモミは約13cmも太くなった(表13).これらの成長により、胸高断面積合計は,2001年の16.7㎡/haから,2017年の34.8㎡/haへと2倍以上になっている.高木層の発達にともない,低木層の構成種(アブラチャン,ナツグミ,マメザクラ)は衰弱し,つる性木本も低木層から亜高木層以上に葉群の中心を移動しつつある.
以上のように,このサイズの風倒ギャップは,下層からの高木生樹種の稚樹の成長を待たず,周囲からの低木の伸長とつる性木本の繁茂を経て,周囲の残存木の樹冠拡大によって閉鎖されることが明らかになった.
(2)種組成の変化
階層ごとの出現種とその優占度の変化を表15に示す(植生調査票は別冊,付表14).高木層の植被率は,2001年の20%から2017年には80%に達した.モミ,イトマキイタヤ,ミズナラのほか,2012年以降は低木層・亜高木層から新出したヤマブドウが加わっている.亜高木層と低木層の植被率は大きく変化せず,アブラチャン,ナツギミ,サンショウバラ,タンナサワフタギ,マメザクラといった低木性樹種,コボタンヅル,ミツバアケビ,ツルウメモドキといったつる性木本が生育していた.亜高木層と低木層の出現種数は2006年が最も多く,それ以降は林冠の閉鎖にともなって減少していた.
ギャップの形成時に樹木の根返りなどは起こらず,土壌は攪乱されていないため,草本層は自然林と近い種組成を保っていた.アズマヤマアザミ,イヌヤマハッカ,ムカゴイラクサなどの高茎草本は,ブナ自然林の調査区よりもむしろ優占度が高いまま維持されていた.これは,ギャップ形成後に低木やつる植物が繁茂したことで,シカが侵入しにくくなり,採食を免れたためと考えられる.
3-2.防鹿柵内外の調査区における5年間の植生変化
3-2-1.防鹿柵内外での種組成と開花植物の違い
防鹿柵の内外に設置した調査区では,2013年以降,年2回ずつ,計10回分の植生調査資料が得られた.表16は,柵内と柵外での出現種を階層別,開花の有無別に示したものである(2017年の植生調査票は別冊,付表15~18).2017年の調査では,柵内で春季82種,秋季75種,柵外で春季80種,秋季71種の植物が記録された.この地域のブナ自然林の林床生草本がほぼ含まれており,シカの採食に対する反応を調査するには好適な場所である.
まず,シカが採食可能な高さ2m以下の第二低木層の変化をみると,柵内外で明瞭な違いがみられる.柵外の調査区では,柵を設置した直後の2013年春の時点で,すでにツクバネウツギとコクサギがわずかに生育しているもみだったが,これらも2015年までに消失し,第二低木層を欠く状態になった.一方,柵内の調査区では,2013年に生育していた低木性樹種は2017年まで継続して出現しただけでなく,2014年からはコクサギが再生し,2016年からはイトマキイタヤの稚樹も加わった.
草本層で柵内だけに新しく出現した種は多くないはないが,2014年以降,ナルコユリ,イヌワラビが,2015年以降,エンレイソウ,ハンショウヅルなどが出現するようになった.しかし,柵外でも2014年からトボシガラ,ギンラン,タニギキョウ,2015年からはギンリョウソウ,ナルコユリ,シオデが新たに出現するなど,柵内だけで種数が増えているわけではない.また柵内外とも,優占度が大きく変化した種もみられなかった.
調査期間中に開花または結実が記録された草本種は53種あり,そのうち34種は柵の内外両方で開花がみられた.柵の内外にかかわらず開花していた種の多くは,草丈が5cm未満でも開花することができる小型の植物か,茎が地表を匍匐する植物であった.草丈が少なくとも10cm以上にならないと開花しない高茎草本は,アズマヤマアザミ,イヌヤマハッカ,ダイコンソウ,ヒロハノハネガヤ,フタリシズカ,ホソバテンナンショウ,ホウチャクソウ,ミズヒキ,ミヤマヤブタバコ,シロヨメナ,トチバニンジンの11種のみであった.また,これらのうち,ダイコンソウ,ヒロハノハネガヤ,フタリシズカ,ホソバテンナンショウ,シロヨメナの5種は,まなびの森付近ではシカの採食痕跡がほとんどみられず,不嗜好植物であると考えられる.
また,柵内外両方に生育していた種のうち,エイザンスミレ,オオイトスゲ,モミジガサ,ナルコユリ,タニギキョウの5種は柵内だけで,ムカゴイラクサ,ヤマシャクヤク(不嗜好植物),ヤマトウバナ,ムラサキケマンの4種は柵外だけで開花がみられた.5年間の追跡調査では,種組成のうえで顕著な変化はみられなかったが,今後も採食圧が続くことで,高茎草本の種子生産が抑制され,徐々に個体群の衰退につながっていくおそれがある一方,小型の植物や不嗜好性植物の優占度が相対的に増加する可能性が高いと考えられる.

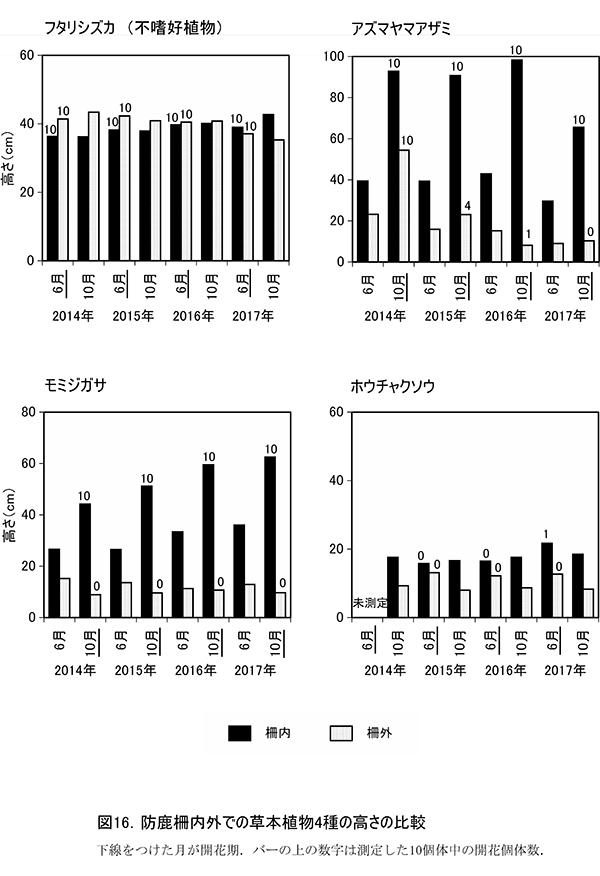
3-2-2.防鹿柵内外での草本種のサイズと開花率の比較
草本種の個体サイズや開花・結実に及ぼすシカの採食の影響を詳しく調べるため,2014年から代表的な草本種7種について,春季と秋季に高さを測定した.このうち,毎年10個体が測定でき,柵内外での比較が可能であったフタリシズカ,アズマヤマアザミ,モミジガサ,ホウチャクソウの4種について,高さの平均値と開花個体数の変化を図16に示した(各個体の測定値は別冊,付表19).
フタリシズカは,春に茎頂に4枚の葉を展開し,その上に数本の花序をつける.花をつけた後はそれ以上伸長しないため,高さは6月,10月ともほとんど変化しない.柵内・柵外とも平均で約35~43cmで,4年間でほとんど変わっていなかった.また,6月には計測したすべての個体が花序をつけていた.このことから,柵外であっても開花を阻害するような採食影響は受けていないといえる.ただし,2017年の計測結果では,柵内と柵外の高さが前年までと逆転しており,不嗜好性植物であっても,これまで若干の影響を受けていた可能性が示唆された.
アズマヤマアザミは,春から秋にかけて茎を伸ばしながら,順次葉を展開し,葉腋に花をつけていく草本である.2016年までは,柵内では6月に40cm程度,10月に1m近くに成長していた.しかし,柵外では6月,10月とも柵内よりも高さが低く,特に10月は柵内に比べて大幅にサイズが小さくなっていた.2016年には,10月の高さが6月よりも低く,わずか8cmまで低下しており,夏の間にシカの採食を受けたことがわかる.開花個体数も,柵内では2017年まで毎年10個体すべてが開花していたが,柵外では2015年に4個体,2016年には1個体と急激に減少し,2017年には開花個体はまったくみられなくなった.2017年では柵内の10月の高さが,前年に比べてかなり低くなっていたが,この原因については明らかでない.
モミジガサは,6月頃までにほとんどの葉を展開し,秋に茎の頂部に花をつける草本である.柵内での10月の高さは年ごとに増加し,2014年の44.2cmから,2016年には62.5cmと約18cmも大きくなった.開花数も2014年以降,10個体を維持していた.一方,柵外では高さは増加せず,6月よりも10月のほうが低くなる傾向もそのままであった.開花も2017年まで,まったくみられなかった.モミジガサは20cm以下の個体サイズでは開花できず,さらに夏の間に茎を採食されてしまうと新しい茎を再生できないため,アズマヤマアザミよりも影響を受けやすいと考えられる.
ホウチャクソウは,春季にすべての葉を展開し,6月頃までに開花する植物である.モミジガサと同様,柵外では6月よりも10月の高さが低くなっており,シカに採食されていることがわかる.柵内でもまだほとんど高さの回復はみられないが,2017年には1個体だけ開花個体が確認された.
以上のように,柵の設置に対する草本種の反応は,種によってさまざまであった.これらの反応の違いからは,それぞれの種が柵の設置までに受けてきた採食圧の影響と,柵によって保護されたときの回復力の違いを読み取ることができる.柵内でのサイズや開花率が変化せず,柵外でのサイズの低下が大きかったアズマヤマアザミは,柵を設置した2014年の時点では,採食圧の影響がほとんど現れていなかったと考えられる.この時点では,アズマヤマアザミはあまり採食されていなかったか,食べられていてもサイズや開花率に影響を及ぼすほどではなかったが,その後も採食圧がかかり続けた柵外では,この4年間で急速に成長の抑制と開花率の低下が生じたといえる.つまり,2013年の柵設置のタイミングは,アズマヤマアザミにとっては,採食の影響が出始める直前であったと考えられるが,2017年の時点では,アズマヤマアザミが開花して種子繁殖を行うことができる採食影響の限度を超えてしまったことになる.
対照的に,モミジガサは柵内のサイズや開花率が大幅に回復している.この種は,柵を設置した2014年までに,繰り返し採食されたことによる地下の貯蔵器官の消耗が激しく,地上部が開花不能なサイズにまで小さくなっていたが,柵で保護されたことによって回復したとみなされる.柵外では,いまでも採食圧がかかり続けているために,高さや開花率は元のままである.モミジガサは,柵設置の時点ですでに大きな採食影響を受けていたものの,採食圧から解放されれば,すみやかに回復できる程度のダメージであったと考えられる.
また,柵設置後もサイズや開花に変化が少ないホウチャクソウは,モミジガサに比べて採食の影響が強く,回復に時間がかかるのかもしれない.これらの結果から,①柵を設置した以外の場所では採食圧がかかり続けており,これまで影響を受けていなかった植物にも影響が出ていること,②柵の設置によって一部の植物は本来の生育状態を回復したが,4年程度では回復できないダメージをすでに被っている種もあること,が明らかになった.現在まで,柵の有無によって種組成レベルの違いは生じていないが,将来的にはシカが好む植物の個体の小型化や繁殖率の低下,不嗜好植物が相対的に有利になることなどによって,種組成にも違いが生じてくることが予想される.


「まなびの森」の20年の活動をまとめたPDFです
![]()