








































モニタリング活動
植生調査 2018年度報告書
1.調査の目的
富士山南麓では1996年(平成8年)9月の台風17号により大規模な風倒被害が発生し,壮齢ヒノキ林を中心に約90,000㎥が被害を受けた.「富士山まなびの森」では,この風倒被害跡地に広葉樹を中心とした在来樹種を植栽することによって,地域本来の自然林を再生することを目指している.風倒木の処理が行われた後,1998年から2002年にかけて,敷地内の約31haにおよそ32,000本の樹木が植栽された.「富士山まなびの森」では,できるだけ人の手を加えずに自然の遷移の進行を生かして森林再生を行う方針が示されているが,針葉樹人工林の跡地に大規模に自然林を復元した事例は過去になく,自然林再生のための管理・施業方法に関する情報はほとんどない.また,1990年代以降,周辺ではニホンジカが急増しており,シカによる高い採食圧のもとで,どのような遷移が起こるかについても情報が必要である.そこで,「富士山まなびの森」実行委員会では,2000年以降,敷地内にいくつかの固定調査区を設置し,植栽地の森林再生過程のモニタリング調査を実施することとした.
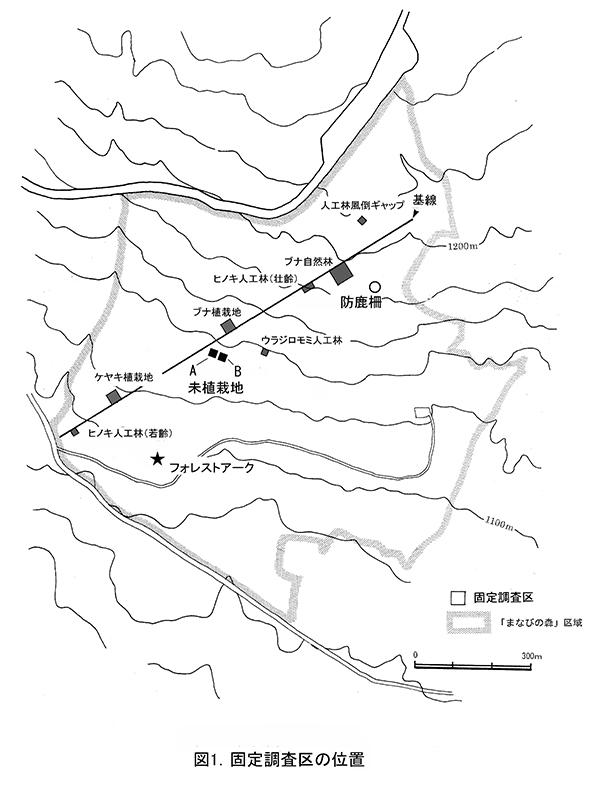
設置された調査区は図1に示す9ヶ所である.植栽地2ヶ所のほか,植栽を行わなかった未植栽地1ヶ所,復元目標としてのブナ自然林や比較対象となる人工林など5ヶ所 に固定調査区を設置した.2013年からは,ブナ自然林内に防鹿柵を設置し,その内外での林床植生の変化についての追跡調査も行っている.
2018年度は,これらのうち,自然の遷移の進行による天然更新の可能性を明らかにするために,あえて樹木の植栽を行わなかった未植栽地の調査区についての追跡調査を行った.未植栽地の調査区は2002年に設置され,その後2005年,2007-2008年,2012年,2015年に追跡調査が行われており,6回目の調査となる.本報告では,過去5回の調査結果と合わせて,調査開始から16年間の森林再生状況についてまとめた.
2.調査地と調査方法
調査区は,富士山国有林166林班わ小班のウラジロモミ人工林に隣接した,標高約1130m,傾斜約3°の南西向き緩斜面にある.基線の東側に10m☓10mの調査区が2つ設置してある(図1).西側の基線寄りのものを調査区A,東側のものを調査区Bとした.この付近では,風倒木の集材後に残った根株や枝条がところどころに集積・放置されているが,調査区は枝条の集積箇所を避けて設置した.なお,設置当初はシカの採食の有無による森林再生の状況を比較することを意図して,調査区を園芸用支柱による柵とネットで囲んだ(調査区A)が,シカの立ち入りを防ぐことができなかったため,シカの採食の有無による比較は行っていない.
各調査区において,2018年10月に以下の調査をおこなった.調査方法は過去の調査結果と比較するために,前回までの調査方法を踏襲した.
①毎木調査
調査区内に生育している樹木について,樹高および直径(根元直径と胸高直径)を測定した.高木性樹種については樹高0.3m以上,低木性樹種については樹高1.3m以上の個体を調査対象とした.樹高は検測棹をもちいてcm単位で測定し,直径は直径割巻尺またはノギスをもちいて㎜単位で計測した.直径については,調査開始時は樹木のサイズが小さかったため根元直径(D0)を測定していたが,樹木の成長にしたがって根張りの影響が大きくなってきたため,2015年の調査時からは胸高直径(高さ1.3mでの直径,DBH)も同時に測定している.
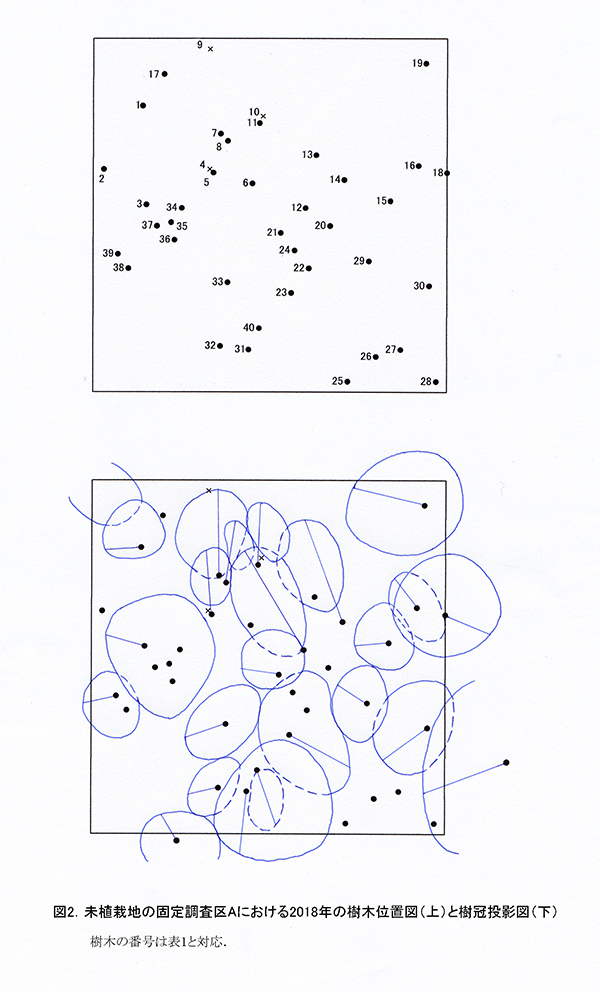
測定した個体には,識別のためにナンバーテープをつけ,樹木の位置図を作成した.また,樹高6m以上の個体については,その樹冠の広がりを図示した樹冠投影図を作成した.
②植生調査
植物社会学的な方法による植生調査をおこなった.これは,出現するすべての植物を階層別にリストアップして,それぞれの種に量的評価(優占度および群度)を与えるものである.優占度は種の生育量,群度は種の生育状態を示す.優占度,群度の判定基準はBraun-Blanquet(1964)にしたがい,優占度はr,+,1~5の7段階,群度は1~5の5段階で記録した.優占度,群度の判定基準は以下の通りである.
【優占度】
| r | : | 単独で生育 |
| + | : | まばらに生育し被度はごく小さい |
| 1 | : | 個体数は多いが被度は小さい(1/10以下) |
| 2 | : | 被度が1/10~1/4 |
| 3 | : | 被度が1/4~1/2 |
| 4 | : | 被度が1/2~3/4 |
| 5 | : | 被度が3/4以上 |
【群度】
| 1 | : | 単独で生育 |
| 2 | : | 小群状または束状に生育 |
| 3 | : | 斑状またはクッション状に生育 |
| 4 | : | 大きな斑状,または穴のあいたカーペット状に生育 |
| 5 | : | 一面に生育 |
3.調査結果
(1)調査区Aの現況と変化
①樹種構成と空間分布
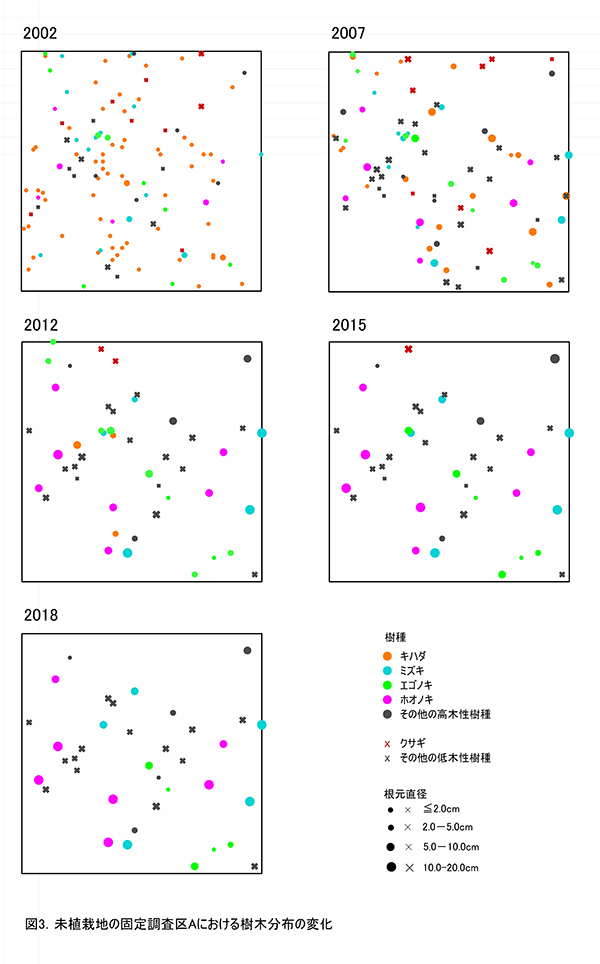
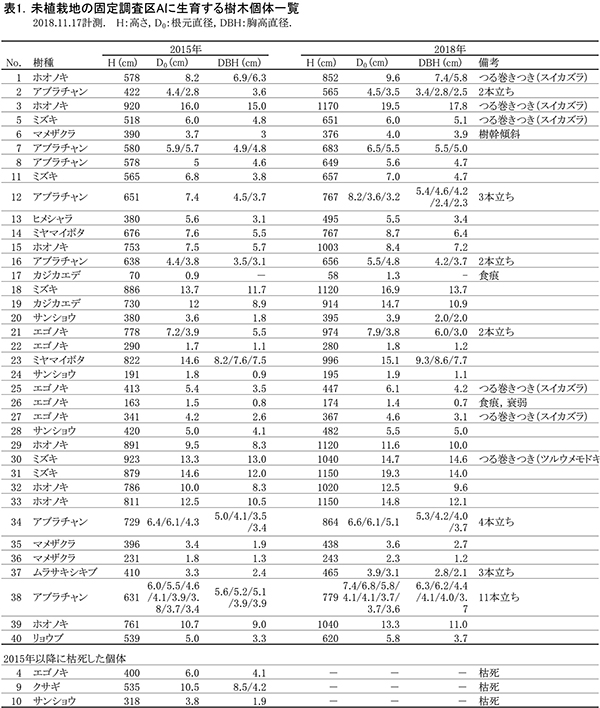
調査区内に生育している樹木の樹高と直径を前回(2015年)の測定値とともに表1に示す.それらの位置および樹冠投影図を図2に示す.また,樹種別の個体数と樹高・直径の経年変化を表2に,樹種別の樹木分布の変化を図3に示す.
2018年現在,調査区Aには高木性樹種6種21本,低木性樹種5種16本の計37本が生育していた.調査を開始した2002年には10種98本の高木性樹種が生育していたが,2007年の12種55本,2012年の9種29本,2015年の6種22本を経て,現在の6種21本となった(表2).樹種別ではホオノキが7本で最も多く,次いでエゴノキとミズキが5本であった.その他,カジカエデ(2本),ヒメシャラ(1本),リョウブ(1本)がみられた.過去3年間で枯死したのはエゴノキ1本のみであった.今回,新たに出現した高木性樹種はなかった.キハダは2002年には63本が生育していたが,2015年までにすべて枯死した.2012年まで生育していたチドリノキとブナも,2015年までに枯死していた.
低木性樹種のうち高さ1.3m以上の個体は5種16本で,前回の6種18本から2本減少した(表2).樹種別では,アブラチャンが7本で最も多く,次いでサンショウとマメザクラが3本であった.その他,ミヤマイボタ(2本),ムラサキシキブ(1本)が見られた.先駆的な性質が強いクサギは,2002年に9本あったが,2018年までに最後の1本が枯死した.低木性樹種でも,今回新たに出現した樹種はなかった.
樹木の分布をみると,ホオノキ,ミズキを主とする根元直径10cm以上の樹木は,ほぼ均等に分布していた(図3).ホオノキとミズキはすべての個体が樹高8mを超えており,これらが林冠を形成しつつある.成長のよいものでは,根元直径が20cm近くに達した個体(No.3,31)もあった.また,低木性樹種でも樹高9mを超えるミヤマイボタ(No.23)もみられた.しかし,互いの樹冠はまだ接しておらず,林冠の閉鎖にはいたっていない(図2).エゴノキはつる植物の巻きつきの影響を受けて成長が悪くなっており,林冠の形成に加わっているのは1本(No.21)のみであった.これら高木性樹種の下層には,複数の幹をもつ高さ6~8mのアブラチャン,ミヤマイボタが樹冠を広げていた.
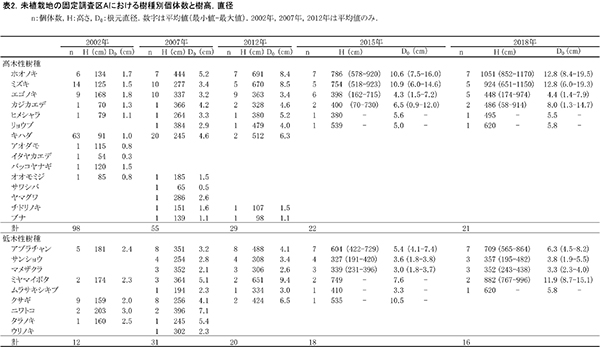
②樹木の成長と階層構造
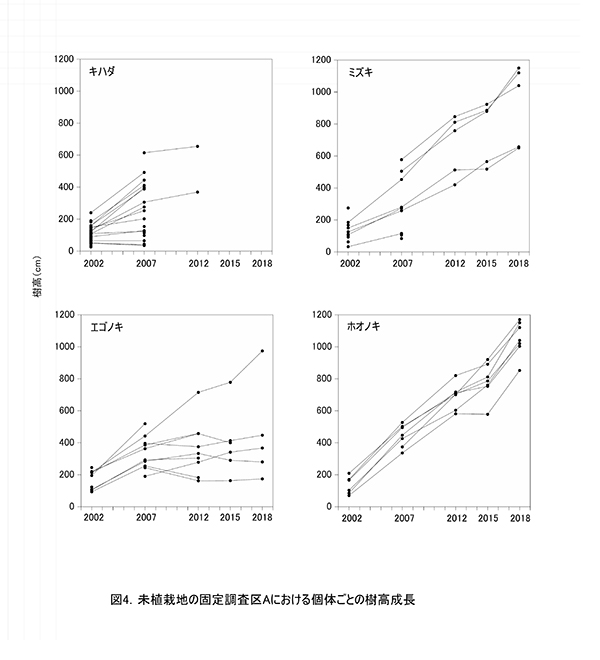
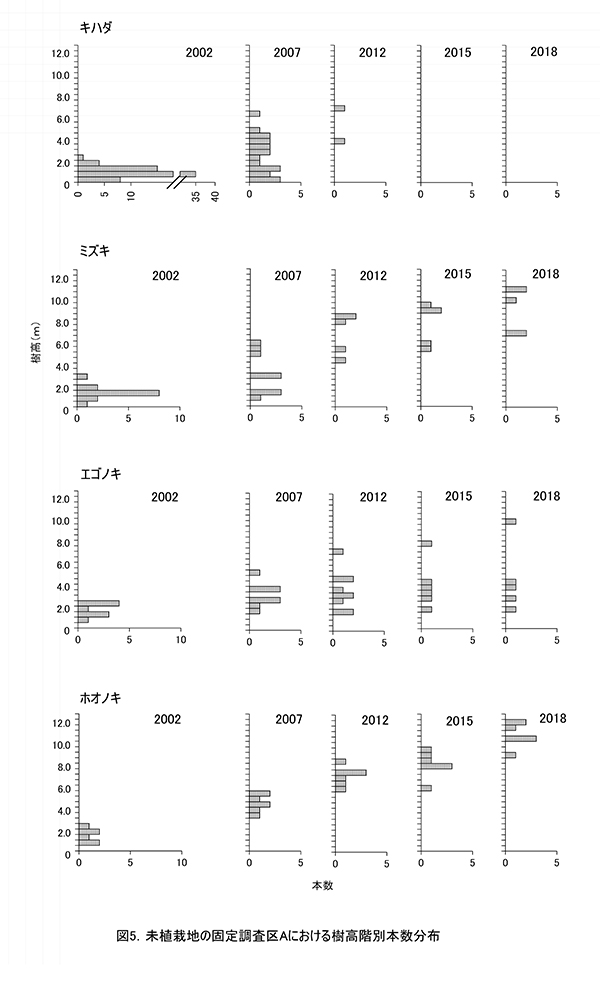
調査区内に生育する高木性樹種のうち,キハダ,ミズキ,エゴノキ,ホオノキの個体ごとの樹高の変化を図4に,これら4種の樹高階級別本数分布の変化を図5示す.さらに図6には,低木性樹種も含めた樹高1.3m以上の全個体を樹高が大きい順に並べた樹高順位曲線を示した.
個体ごとの樹高成長をみると,樹種によって成長過程に特徴があることがわかる(図4).調査開始当初,多数の個体が存在していたキハダは,2007年までに樹高5mを超えられなかった個体はほとんど枯死していた.2005年の植生調査資料によると,このとき低木層は高さ6mで植被率60%,草本層は高さ1.8mで植被率95%に達していた.そのため,光要求性が高いキハダは,他の低木性樹種や草本層で繁茂していたススキとの競争によって大半が枯死したものと考えられる.残った2個体も2012年までには低木層の高さを超えられずに枯死していた.
これに対して,ミズキは2007年にススキが優占する草本層の高さを超えられなかった3個体は枯死したものの,高さ2m以上のものはその後も生存し続けていた.ホオノキも同様に,2007年に樹高3~5mの範囲にあったものが,その後もすべて生存していた.これら2種は2012年から2015年にかけてやや成長速度が低下したが,それ以降生長量が回復し,樹高10mを超える個体も現れた.ミズキとホオノキは,キハダに比べると耐陰性があり,クサギやアブラチャンなどの低木性樹種と葉群が重なる状況下でも生存できたことがわかる.
エゴノキは,2007年に4mを超えていた1本だけがその後も順調に成長し,4m未満の個体は樹高成長が頭打ちになっていた.エゴノキは,ススキや低木性樹種に被圧された状態でもすぐには枯死せず,10年以上も生存し続けるだけの耐陰性をもつことがわかる.2018年にはススキや低木性樹種による被圧からは開放されているが,現存する個体はスイカズラなどのつる植物に巻きつかれており,衰弱している.そのため,現時点で樹高4m未満の個体が林冠に加わっていくことは難しいと考えられる.
樹高順位曲線(図6)の経年変化をみると,2002年には上位にエゴノキが多かったが,次第にホオノキやミズキが上位に進出し,2012年にはほぼこの2種が上層を占めていたことがわかる.2018年には,樹高8m以上の14本のうち11本をホオノキとミズキが占めていた.樹高順位曲線の傾きの変化は階層構造の発達を意味する.2018年の樹高順位曲線では,7-8m付近と5m付近に傾きの不連続があり,林冠を構成する樹木と,低木層を構成する樹木が分かれつつある.この図からも,エゴノキの多くが低木層に取り残されていることがわかる.
③種組成
表3に調査区Aの植生調査票を示す.林冠を形成しつつある高さ7~10mの層を亜高木層とした.7m以下の低木層の中でも,下層に被圧される樹木がでてきたため,低木層を二つに分け,下層を第二低木層とした.亜高木層に5種,第一低木層に12種,第二低木層に6種,草本層に59種,重複を除くと計70種の植物が記録された.出現種数は前回(2015年)の80種から10種減少していた.
亜高木層の植被率は前回の60%から70%に増加し,ホオノキの優占度が最も高かった.そのほか,ミズキ,エゴノキ,カジカエデ,ミヤマイボタが混生していた.第一低木層の植被率は前回と変わらず50%であった.アブラチャンが最も高い優占度を占め,ミズキ,ヒメシャラ,リョウブなどの高木性樹種のほかに,ガマズミ,サンショウ,ムラサキシキブなどの低木性樹種が生育していた.また,つる植物のスイカズラやクマヤナギが低木層の樹冠上に広がっていた.
草本層の植被率は40%で,前回の85%でから大きく減少した.これは,8月の台風による降雨で,調査区内に土砂が流れ込み,林床植生が埋没したためである.最も優占度が高かったのは,シカの不嗜好植物であるシロヨメナで,優占度3で生育していた.同じくシカの不嗜好植物であるアシボソ,フタリシズカも優占度1で生育していた.前回から新たに出現した種には,ツルシロカネソウ,モミジガサ,クモキリソウなどの林床生草本,キツネノボタン,オトコエシなど攪乱された立地に生育する草本,クマヤナギ,ミツバアケビなどのつる性木本 を含む11種であった.一方,前回は優占度1であったススキが完全に消失した.ススキは最盛期の2007年には優占度3まで増加したが,その後減少し続け,林床の優占種はシロヨメナに置き換わった.その他,アオホオズキ,エイザンスミレなどの林床生草本,コアカソ,ミツバウツギなどの低木,シダ植物のハリガネシダ,ゲジゲジシダなど計20種がみられなくなった.土砂の流入の影響が大きいため,種の消失については,遷移の進行によるものかどうかは一概に判断できないが,少なくともススキの消失は,林冠の発達にともなう林床の明るさの低下によるものと考えられる.
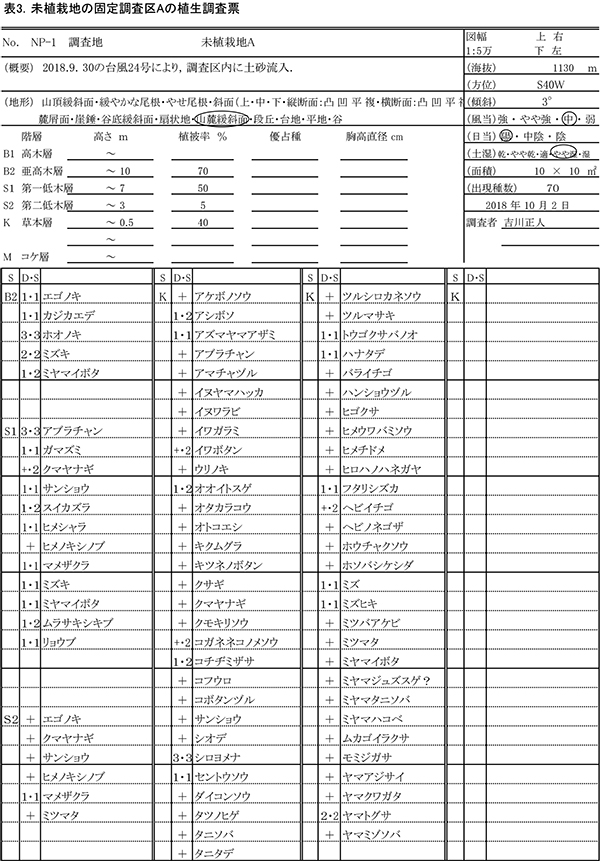
(2)調査区Bの現況と変化
①樹種構成と空間分布
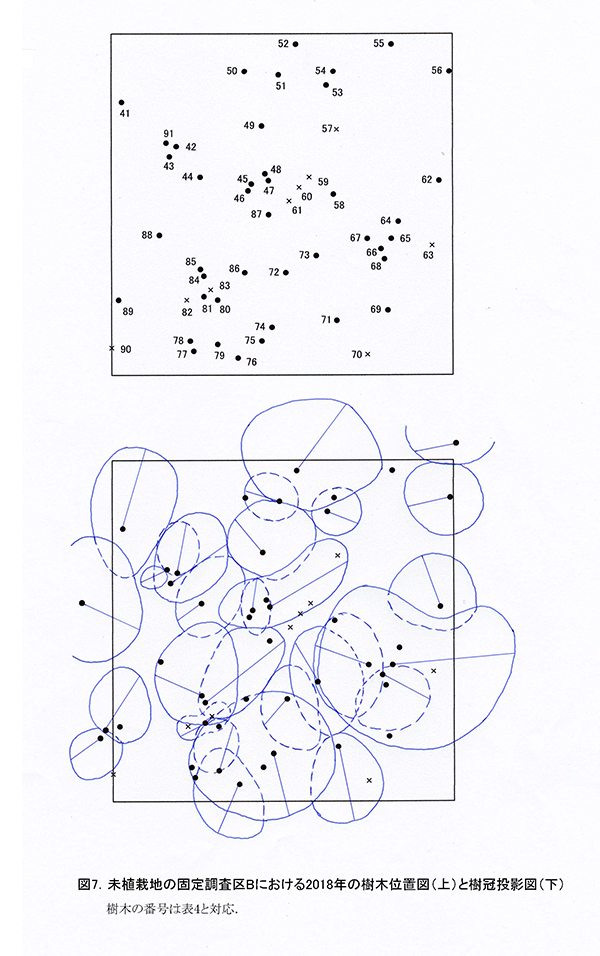
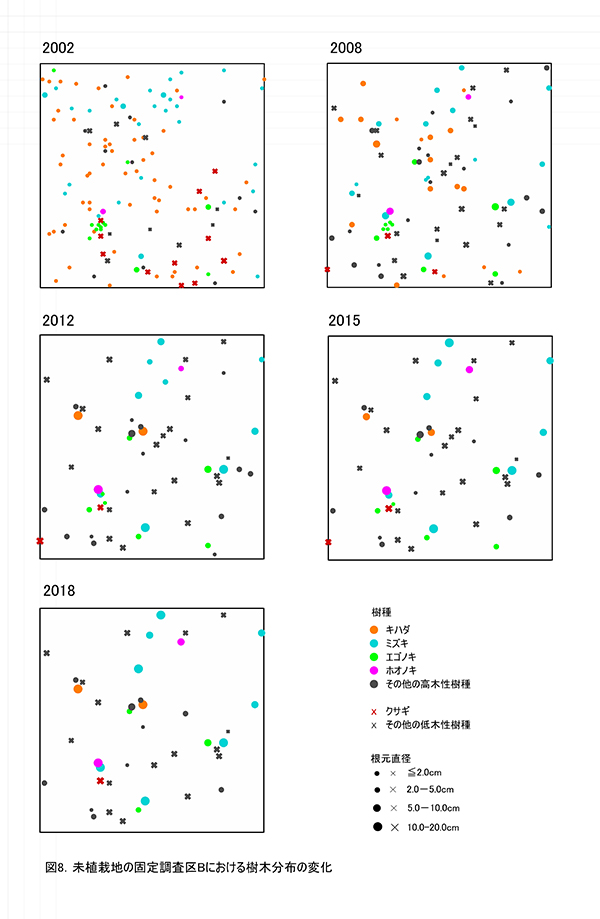
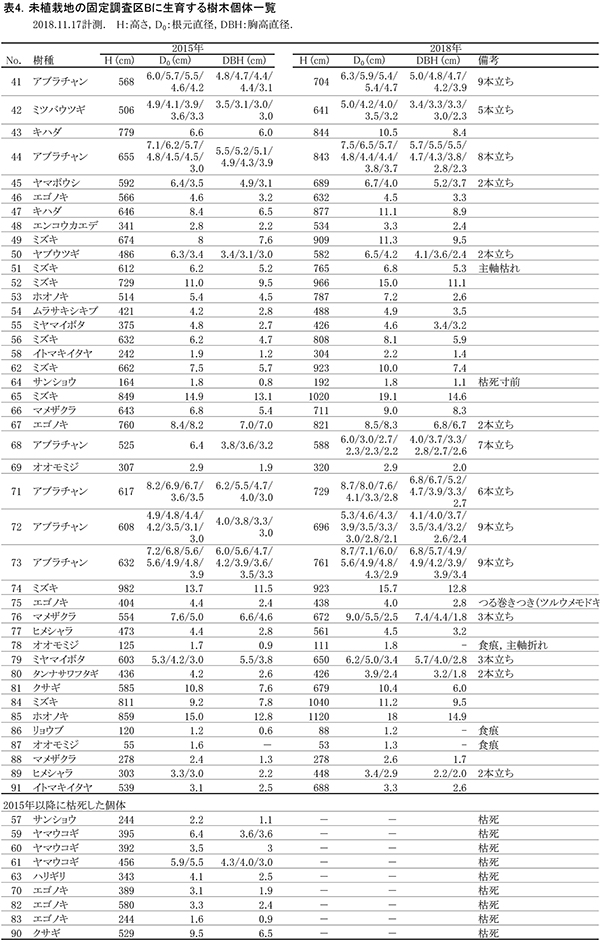
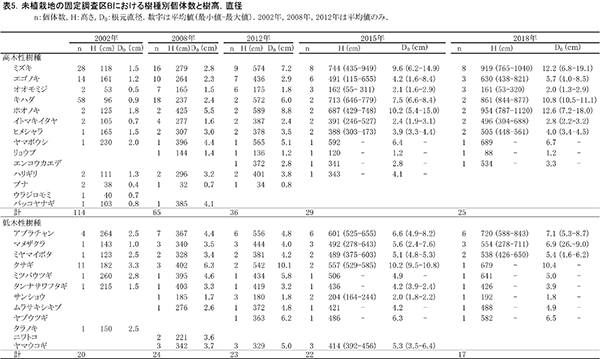
調査区内に生育している樹木の樹高と直径を前回(2015年)の測定値とともに表4に示す.それらの位置および樹冠投影図を図7に示す.また,樹種別の個体数と樹高・直径の経年変化を表5に,樹種別の樹木分布の変化を図8に示す.
2018年現在,調査区Bには高木性樹種10種25本,低木性樹種9種17本の計42本が生育していた.調査を開始した2002年には12種114本の高木性樹種が生育していたが,2007年の12種65本,2012年の12種36本,2015年の11種29本を経て,現在の9種25本となった(表5).樹種別ではミズキが8本で最も多く,次いでエゴノキとオオモミジが3本であった.その他,ホオノキ(2本),キハダ(2本),イトマキイタヤ(2本),ヒメシャラ(2本)などがみられ,調査区Aよりも構成樹種が多様である.過去3年間で枯死したのはエゴノキ3本とハリギリ1本であった.今回,新たに出現した高木性樹種はなかった.この調査区でも,2002年の調査開始当初はキハダの個体数が最も多く58本が生育していたが,現在では2本が残るのみである.
低木性樹種のうち高さ1.3m以上の個体は9種17本で,前回の9種22本から5本減少した(表5).樹種別では,アブラチャンが6本で最も多く,次いでマメザクラが3本,ミヤマイボタが2本であった.先駆的な性質が強いクサギは,2002年には11本あったが,2018年には1本が残るのみであった.低木性樹種でも,今回新たに出現した樹種はなかった.
樹木の分布をみると,根元直径10cm以上のミズキ,キハダ,ホオノキがほぼ均等に分布していた(図8).これらは樹高8mを超えるものが多く,円形の樹冠を広げて林冠を形成しつつあるが,互いの樹冠はまだ接しておらず,林冠の閉鎖にはいたっていない(図7).No.65のミズキは最も成長がよく,根元直径が20cm近くに達し,樹冠面積も大きかった.一方で,No.43と47のキハダの樹冠は,やや偏った形状を示していた(表5,図7).下層には複数の幹をもつ高さ6~8mのアブラチャンが,高木性樹種の樹冠の隙間を埋めるように樹冠を広げていた.
②樹木の成長と階層構造
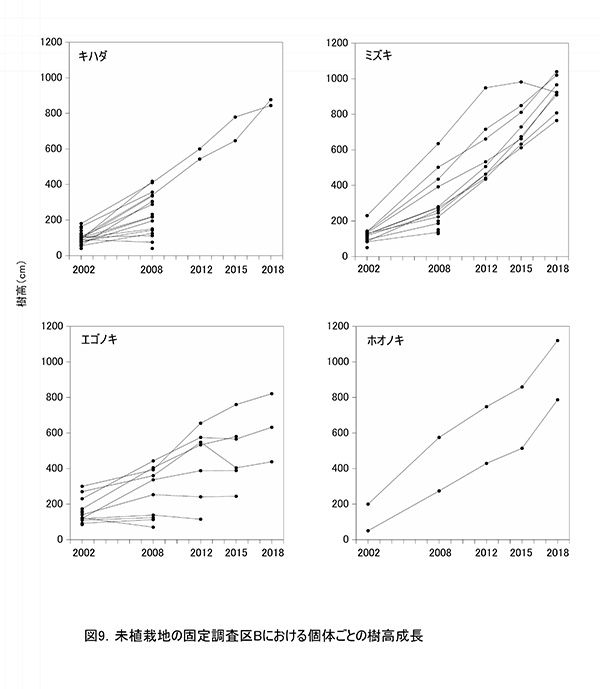
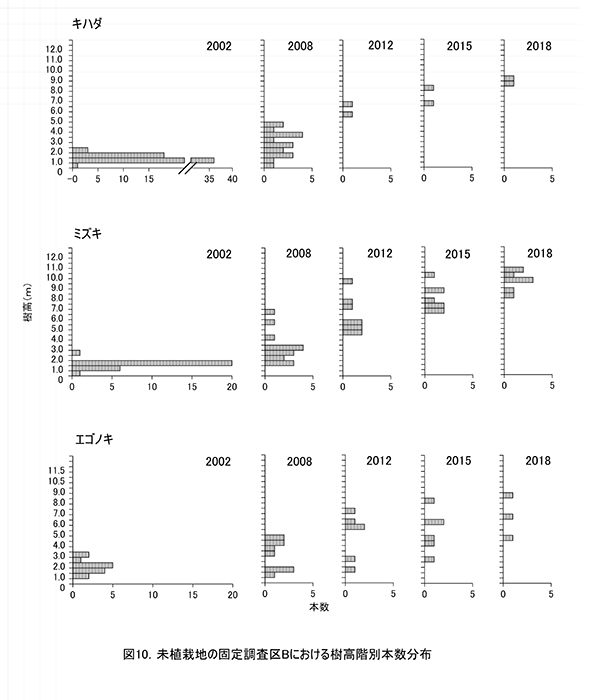
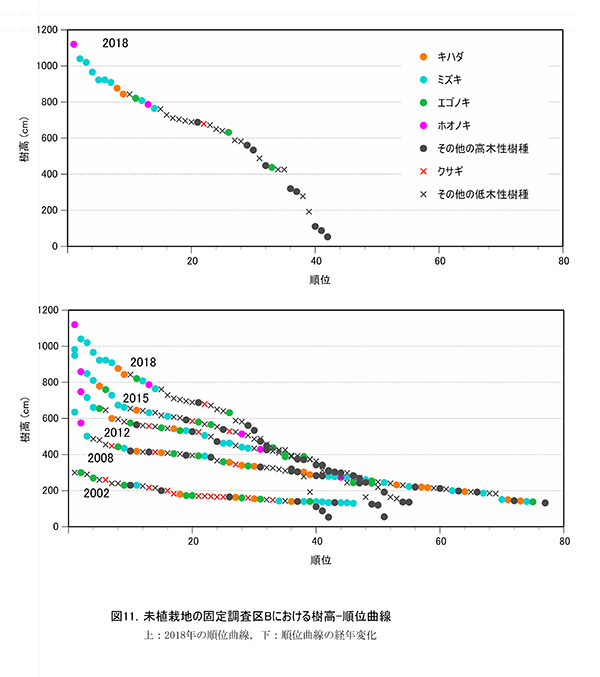
調査区内に生育する高木性樹種のうち,キハダ,ミズキ,エゴノキ,ホオノキの個体ごとの樹高の変化を図9に,これら4種の樹高階級別本数分布の変化を図10示す.さらに図11には,低木性樹種も含めた樹高1.3m以上の全個体を樹高が大きい順に並べた樹高順位曲線を示した.
個体ごとの樹高成長をみると,調査区Aと同様に,樹種によって特徴的な成長パターンがあることがわかる(図9).調査開始当初,多数の個体が存在していたキハダは,2008年までに樹高4mを超えられなかった個体は1本を除いてすべて枯死していた.2005年の植生調査資料によると,このとき低木層は高さ5mで植被率80%に及び,多数の高木性樹種の稚樹のほか,クサギ,アブラチャン,ミツバウツギなどの低木が繁茂していた.さらに草本層は高さ1.8mで植被率95%に達し,ススキやモミジイチゴが密な植被を形成していた.そのため,この頃までに低木層の高さを超えられなかったキハダの稚樹は,被圧によって大半が枯死したものと考えられる.2012年に低木層から抜け出していた2本は,現在でも生存している.
これに対して,ミズキは2008年に草本層の高さを超えられなかった個体は枯死したものの,高さ2m以上のものはその後も生存し続けていた.ホオノキは2本とも,2008年には樹高2mを超えており,その後も生存していた.これら2種は2008年以降ほぼ一定の速度で樹高成長を続け,2018年には樹高10mを超える個体も現れた.ただし,ミズキ1個体だけは梢端に枯れが生じ,2012年以降の成長が頭打ちになっていた.このように,ミズキとホオノキは,高さ2m未満の段階での耐陰性がキハダよりも強かったと考えられる.
エゴノキは,2008年までに4mを超えられなかった個体も,いくつかが生存し続けていたが,成長が頭打ちになった個体は2015年までに順次枯死し,2018年には樹高4m以上の3本が残るのみとなった.やはりある程度被圧に耐える能力はあるが,低木層を抜けるのに時間を要するため,林冠の形成に加われる個体は少なくなると考えられた.
樹高順位曲線(図11)の経年変化をみると,2002年にはエゴノキと低木性樹種が上位を占めていたが,次第にホオノキ,ミズキ,キハダが上位に進出してきたことがわかる.2018年には,樹高8m以上の12本のうち7本をミズキが占めていた.アブラチャンなどの低木性樹種は,樹高7m付近が上限となっており,林冠層と低木層の分離が進行していることが読み取れる.また,カエデ類など遷移後期の高木性樹種は,アブラチャンなどよりも低い樹高6m以下にとどまっている個体が多かった.
③種組成
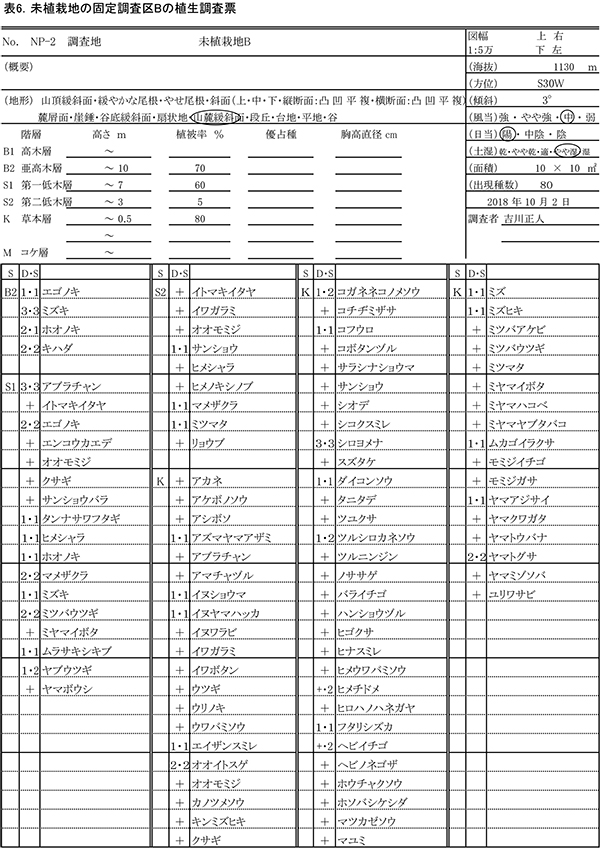
表6に調査区Bの植生調査票を示す.林冠を形成しつつある高さ7~10mの層を亜高木層とした.7m以下の低木層の中でも,下層に被圧される樹木がでてきたため,低木層を二つに分け,3m以下を第二低木層とした.亜高木層に4種,第一低木層に14種,第二低木層に9種,草本層に67種,重複を除くと計80種の植物が記録された.出現種数は前回(2015年)の92種から12種減少していた.
亜高木層の植被率は前回の50%から70%に増加し,ミズキの優占度が最も高かった.そのほか,ホオノキ,キハダ,エゴノキが混生していた.第一低木層の植被率は50%から60%に増加していた.アブラチャンが最も高い優占度を占め,エゴノキ,ミズキ,ホオノキ,ヒメシャラなど高木性樹種のほかに,マメザクラ,ミツバウツギ,ムラサキシキブなどの低木性樹種が生育していた.先駆的性質が強い低木では,ヤマウコギが枯死したが,ヤブウツギやサンショウバラはまだ残存していた.
調査区Bは大雨による土砂の流入を受けていなかったため,草本層の植被率は80%で前回と同じであった.最も優占度が高かったのは,シカの不嗜好植物であるシロヨメナであったが,優占度は4から3に低下した.同じくシカの不嗜好植物であるフタリシズカ,ダイコンソウも優占度1で生育していた.そのほか,アズマヤマアザミ,イヌショウマ,イヌヤマハッカなどの高茎草本,オオイトスゲ,エイザンスミレ,コガネネコノメソウ,ムカゴイラクサ,ヤマトグサなどの小型の草本が優占度1以上で生育していた.前回から新たに出現した種には,シコクスミレ,ヤマクワガタ,イワボタン,ユリワサビなど林床に生育する草本を含む11種であった.一方,アズマレイジンソウ,チダケサシ,フユノハワワラビといった草本や,ガマズミ,ツノハシバミ,メギなどの木本を含む14種が確認できなかった.前回は優占度+であったススキも完全に消失した.ススキは調査開始当初の2002年には優占度3であったが,その後減少し続けている.ススキが消失したことで,草本層の高さは0.5mに低下し, 低木層との間に空間ができて,階層構造が明瞭になりつつある.
 調査区A
調査区A
 調査区B
調査区B
写真1.未植栽地の固定調査区設置前の状況(2000年10月12日撮影)
1996年の風倒被害発生から4年後.シロヨメナ,アズマヤマアザミ,ススキなどの草本が繁茂し,クサギ,ヤブウツギといった先駆性の低木が成長している.高木性樹種の稚樹はまだ目立たない.
 調査区A
調査区A
 調査区B
調査区B
写真2.未植栽地の固定調査区(2012年5月31日撮影) 風倒被害から16年後.クサギなどの先駆性低木は衰退し,アブラチャンなどが高さ4~5mの低木層を形成.ホオノキ,ミズキなどの一部の個体が低木層を抜け始める.林床ではススキが衰退し,シロヨメナが優占.
 調査区A
調査区A
 調査区B
調査区B
写真3.未植栽地の固定調査区(2015年10月4日撮影) 低木層がさらに高くなり,林内を楽に歩けるようになる.林床はシロヨメナの優占度が高くなる.
 調査区A
調査区A
 調査区B
調査区B
写真4.未植栽地の固定調査区(2018年10月2日撮影) ホオノキ,ミズキなどが高さ10m前後まで成長し林冠を形成しつつあるが,まだ閉鎖はしていない.調査区Aでは台風による土砂の流入で,林床植生の一部が埋没.


「まなびの森」の20年の活動をまとめたPDFです
![]()